Моногибридным скрещиванием называют такое, при котором родительские формы различаются лишь по одной паре альтернативных или контрастирующих признаков.
Например, отцовское растение несет пурпурные цветки, а материнское — белые, или наоборот.
Перед тем как производить скрещивание, необходимо убедиться в том, что избранные признаки родительских форм являются контрастирующих в ряду поколений, т. е. при самоопылении или близко — родственном скрещивании каждый из избранных признаков стойко наследуется. Родственные организмы, воспроизводящие в ряду поколений одни и те же наследственно константные признаки, принято называть линией.
У растений с гермафродитными цветками при искусственной гибридизации до опыления производят кастрацию цветков материнского растения, удаляя пыльники до того, как они созрели. Однополые женские цветки перекрестноопылителей заблаговременно помещают в изоляторы. В момент созревания рылец на них наносят пыльцу, собранную с цветков отцовского растения.
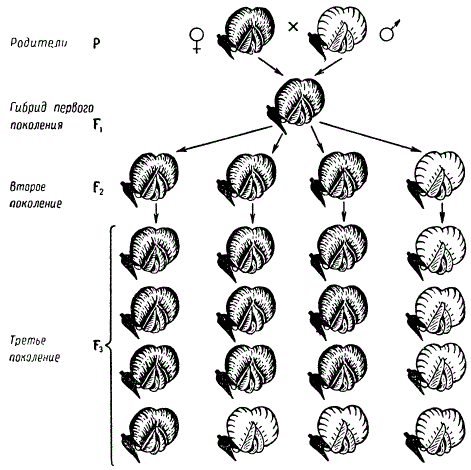
Когда скрещиваются растения, например, гороха, то семена, созревшие в бобе на материнском растении в год скрещивания, являются уже гибридами первого поколения (F1). Из этих посеянных семян вырастут гибридные растения первого поколения, а в бобах этих растений в результате самоопыления разовьются семена с зародышами второго поколения (F2). Если материнское растение имело цветки, например, пурпурные, а отцовское — белые, то цветки гибридных растений F1 оказываются все пурпурными, растения с белой окраской цветков среди них не появляются.
От скрещивания растений, различающихся по окраске семян (желтые и зеленые), на материнском растении гибридные семена в год скрещивания оказываются только желтыми. Если исходные растения отличались по форме семян (гладкие — морщинистые), то гибридные семена F1 на материнском растении оказываются только гладкими.
Следовательно, у гибрида первого поколения из каждой пары альтернативных признаков развивается только один. Второй признак как бы исчезает, не проявляется. Это явление преобладания гибрида признака одного из родителей Г. Мендель назвал доминированием. Признак, проявляющийся у гибрида первого поколения и подавляющий развитие другого признака, был назван доминантным; противоположный, т. е. подавляемый, признак — рецессивным. Мендель так определяет доминантные и рецессивные признаки: «Признаки, которые переходят в гибридные соединения совершенно неизменными или почти неизмененными и тем самым представляют признаки гибридов, будут обозначаться как доминирующие, а те, которые становятся при гибридизации латентными, — как рецессивные».
Закон доминирования — первый закон Менделя — называют также законом единообразия гибридов первого поколения, так как все особи первого поколения имеют одинаковое проявление признака.
Если гибриду первого поколения, полученному, например, от скрещивания двух форм гороха, различающихся по окраске цветка, представляется возможность самоопыляться, то в следующем поколении, т. е. в F2 появляются растения с признаками обоих родителей. Это явление носит название расщепления. В F2 наблюдается расщепление в совершенно определенном количественном соотношении, а именно: в среднем 3/4 от общего числа растений несут пурпурные цветки и лишь 1/4 — белые, т. е. отношение числа растений с доминантным признаком к числу растений с рецессивным признаком оказывается равным 3 : 1. Следовательно, рецессивный признак у гибрида первого поколения не исчез, а был только подавлен и проявился во втором гибридном поколении.
Каждое растение из F2 с белыми цветками при самоопылении в следующих поколениях — F3 и F4 и т. д. дает растения только с белыми цветками. Растения с пурпурными цветками ведут себя иначе. Лишь 1/3 из них при самоопылении дает в F3 и следующих поколениях растения только с пурпурными цветками, а остальные 2/3 вновь дают растения обоих типов в отношении: 3 растения с пурпурными цветками и 1 — с белыми.
Наследование пурпурной и белой окрасок цветов у Pisum sativum
Следовательно, класс растений F3 с доминантным признаком распадается по своим наследственным задаткам в отношении 1 : 2, а все растения второго поколения дают отношение при расщеплении по одной паре наследственных задатков 1 : 2 : 1. Понятие класса здесь и в последующем употребляется в смысле группы потомков, сходных по изучаемому признаку или наследственным задаткам.
Все изложенное в отношении наследования окраски цветка приложимо и к наследованию любой другой пары альтернативных признаков, правда, при определенных условиях, о чем будет сказано ниже.
Так, при изучении наследования гладкой или морщинистой формы семян от 253 гибридных самоопыляющихся растений F1 Менделем было получено в F2 7324 семени, из них гладких — 5474, морщинистых — 1850. Если отношение 3 : 1 является правильным, то при общем числе семян 7324 теоретически следовало ожидать следующее распределение: 1/4 семян (т. е. 7324 Х 1/4 = 1831) должна обладать рецессивным признаком (морщинистые), а 3/4 (т. е. 7324 Х 3/4 = 5493) — доминантным (гладкие). В опытах Менделя были получены цифры, очень близкие к теоретическим.
В другом опыте, где учитывался признак окраски семян (желтые или зеленые), Мендель получил следующее соотношение в F2: из 8023 семян оказалось 6022 желтых и 2001 — зеленое, т. е. опять-таки отношение, очень близкое к 3 : 1.
Однако Мендель неоднократно подчеркивал, что эти отношения отражают лишь средние величины; при малом числе особей количество растений с альтернативными признаками в F2 будет колебаться в силу случайных причин.
Данные свидетельствуют о том, что у отдельных растений имеются очевидные колебания в соотношении классов семян, но в сумме получается отношение, близкое к ожидаемому распределению 3:1. Опытные данные дали очень близкое к этому отношение — 355 : 123.
Итак, проведя моногибридные скрещивания, Мендель установил следующие закономерности наследования.
1. У гибридов первого поколения проявляется только один из пары альтернативных признаков — доминантный, рецессивный же не проявляется. Это явление было названо доминированием, а позднее — первым законом Менделя, или законом единообразия гибридов первого поколения.
2. Во втором поколении гибридов появляются особи как с доминантным признаком, так и с рецессивным, отношение первых ко вторым в среднем равно 3:1. Это явление в 1900 г. Г. де Фриз предложил назвать законом расщепления, а впоследствии оно было названо вторым законом Менделя. Потомки с рецессивным признаком в последующих поколениях при самоопылении остаются константными.
3. Среди 3/4 растений второго поколения с доминантным признаком 2/4 от общего числа потомков оказываются гибридными; при самоопылении они дают в F3 расщепление вновь в отношении 3:1, и только 2/4 остается константной в последующих поколениях, подобно исходным родительским формам и растениям из F2 с рецессивным признаком. Следовательно, в F2 половина растений является гибридными, а половина — «чистыми», константно сохраняющими родительские признаки. Таким образом, потомки гибридов F1 по данным наследственным признакам расщепляются в отношении 1 : 2 : 1. Мендель формулировал это следующим образом: «Гибриды по двум различающимся признакам образуют семена, из которых половина дает вновь гибридную форму, тогда как другая дает растения, которые остаются константными, в равных долях содержат доминирующий и рецессивный признаки».
Как мы видели, в F2 следует различать, во-первых, расщепление по внешнему проявлению признаков, которое выражается отношением, 3: 1, и, во-вторых, по наследственным потенциям, задаткам, выражаемое отношением 1:2:1. Первый тип расщепления называют расщеплением по фенотипу, т. е. по внешнему проявлению признаков, второй тип — по генотипу, т. е. по наследственным задаткам. Термины «фенотип» и «генотип» введены в 1903 г. В. Иоганнсеном.
Под генотипом мы понимаем совокупность наследственных задатков, которыми обладает организм. Фенотипом называют совокупность свойств и признаков организма, которые являются результатом взаимодействия генотипа особи и окружающей среды.
Мендель впервые применил символическое обозначение генотипа, где наследственные факторы, определяющие парные альтернативные признаки, обозначались буквами латинского алфавита. Такую пару альтернативных признаков В. Бэтсон в 1902 г. предложил назвать аллеломорфной парой, а парность признаков — аллеломорфизмом. Б 1926 г. В. Иоганнсен предложил термин «аллеломорфизм» заменить более кратким — «аллелизм», а отдельный фактор одной пары назвать «аллелью». Под терминами «доминантная» или «рецессивная аллель» стали понимать альтернативное состояние одного и того же гена. Этот же смысл был придан и прежним прочно закрепившимся в генетике обозначениям — «доминантный ген» и «рецессивный ген».
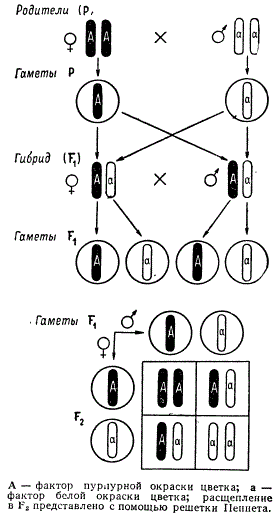
Доминантную аллель, определяющую признак желтой окраски семян, Мендель обозначил заглавной буквой А, рецессивную аллель, определяющую признак зеленой окраски, — строчной буквой а; генотип доминантной формы — АА, а рецессивной — аа; гибрид F1 — Аа. В таком случае потомки гибридов показывают расщепление в F2, соответствующее формуле: 1АА : 2Аа : 1аа.
Эта символика факторов парных признаков, используемая для отображения закономерностей расщепления в потомстве гибридов, является своеобразной «алгеброй» генетики, поскольку вместо буквенных выражений могут быть подставлены соответствующие им различные гены и аллели.
Константные формы АА и аа, которые в последующих поколениях не дают расщепления, В. Бэтсон в 1902 г. предложил назвать гомозиготными, а формы Аа, дающие расщепление, — гетерозиготными. Эти термины широко используются в генетике. Они происходят от термина «зигота», т. е. оплодотворенная яйцеклетка.
Соединение в процессе оплодотворения одинаковых по факторам мужских и женских гамет А и А или а и а дает гомозиготу, или гомозиготную особь АА или аа, а соединение гамет, различающихся по факторам А и а, — гетерозиготу, или гетерозиготную особь Аа.
Как мы видели, у гибридов первого поколения рецессивная аллель а хотя и не проявляется, но и не смешивается с доминантной А, а во втором поколении обе аллели вновь проявляются в «чистом» виде. Такое явление можно объяснить, лишь исходя из допущения, что гибрид первого поколения Аа образует не гибридные, а чистые гаметы, при этом указанные аллели оказываются в разных гаметах. Гаметы, несущие аллели А и а, образуются в равном числе; исходя из этого становится понятным расщепление по генотипу 1:2:1.
Несмешивание аллелей каждой пары альтернативных признаков в гаметах гибридного организма называют явлением чистоты гамет, в основе второго лежит цитологический механизм мейоза.
Мендель обнаружил интересное явление изменения соотношения гомозигот и гетерозигот в ряду последовательных поколений гибрида при самоопылении. Если допустить, что в среднем все растения в ряду поколений имеют одинаковую плодовитость и жизнеспособность и принять во внимание факт расщепления гибридов во втором поколении в определенном числовом отношении (а именно 2 гомозиготных и 2 гетерозиготных растения по данному признаку), то в последующих поколениях при самоопылении всех растений число гомозигот будет увеличиваться, а гетерозигот — уменьшаться.
Для простоты Мендель предположил, что каждое растение при самоопылении дает в каждом поколении четыре семени. В таком случае соотношение гомозигот и гетерозигот по одной паре признаков будет изменяться в ряду поколений. В десятом поколении, по расчету Менделя, на каждые 2048 растений, которые должны возникнуть при взятом коэффициенте размножения, 1023 будут константными — гомозиготными по доминантному признаку, 1023 — гомозиготными по рецессивному признаку и появятся только 2 гибридные, т. е. гетерозиготные формы. Эти расчеты Менделя, как мы убедимся в последующем, имеют значение для понимания генетических основ селекции и динамики генов в популяции.
До сих пор мы говорили об одной паре альтернативных признаков, один из которых является доминантным, другой — рецессивным. Мендель исследовал у гороха семь пар таких признаков:
1) семена гладкие и морщинистые, или угловатые, 2) семядоли желтые и зеленые, 3) окраска семенной кожуры серо-коричневая и белая, 4) форма боба выпуклая и с перетяжками, 5) окраска зрелого боба желтая и зеленая, 6) расположение цветков пазушное и верхушечное, 7) стебель растения высокий и низкий. По каждой из этих семи пар признаков в отдельности в F2 наблюдалось расщепление по фенотипу в среднем в отношении 3:1. Выражая это в процентах, можно сказать, что в F2 было около 75% растений с доминантным и около 25% с рецессивным признаками, или доминантных гомозигот — 25%, гетерозигот — 50% и рецессивных гомозигот — 25%.
Истинная природа парности признаков Менделю осталась неизвестной. Он предполагал, что половые клетки несут по одному наследственному задатку, которые попарно соединяются при оплодотворении. Теперь эти задатки, или факторы, переносимые половыми клетками, называют генами. Под термином «ген» пока мы будем понимать единицу наследственности, определяющую развитие отдельного признака, или свойства, организма. По мере углубления анализа явлений наследственности и механизма наследования наши представления о природе гена будут расширяться. Гены, как мы указали, определяют развитие признаков организма и должны быть связаны с материальными структурами половых клеток. Очевидно, во времена Менделя на этот счет можно было высказывать лишь догадки, поскольку строение и развитие половых клеток не было еще изучено.
Чем же определяется парность генов, чистота гамет и строгое распределение генов в потомстве, обусловливающее расщепление в определенном числовом отношении? После того как мы познакомились в предыдущих главах с развитием половых клеток и формированием гамет, нам несложно связать наблюдавшееся Менделем явление расщепления признаков с поведением хромосом: их парностью, расхождением гомологичных хромосом в мейозе и воссоединением их в процессе оплодотворения.
Допустим, что в соматических клетках растения имеется всего одна пара гомологичных хромосом, а ген, определяющий признак пурпурной окраски цветка, обозначаемый А, находится в каждой из этих хромосом у родительского растения. Тогда соматические клетки гомозиготного растения, обладающие доминантным геном окраски цветка, должны нести две доминантные аллели АА, поскольку в соматических клетках каждая из гомологичных хромосом представлена в двойном количестве. Соответственно клетки другого родительского растения с белыми цветками имеют в гомозиготе рецессивную аллель белой окраски, т. е. аа.
В результате мейоза в каждой гамете число хромосом уменьшается в два раза и остается только одна хромосома из пары. Следовательно, ген, находящийся в этой хромосоме, в гамете представлен в виде аллелей А или а. В результате оплодотворения в гибридной зиготе восстанавливается парность хромосом, и формула гибрида будет точно такой, как ее написал Мендель — Аа. При развитии половых клеток в гибридном организме в мейозе хромосомы данной пары разойдутся в разные дочерние клетки. Тогда мужские и женские гаметы будут нести по одной из аллелей гена: А или а. Такие гаметы, как мужские, так и женские, будут образовываться в равном числе. При оплодотворении типов могут соединяться с равной вероятностью образуется четыре типа зигот.
Поведение пары гомологичных хромосом при моногибридном скрещивании
Для облегчения расчета сочетаний разных типов гамет английский генетик Р. Пеннет предложил производить запись в виде решетки, которая и вошла в литературу как решетка Пеннета. Влево от решетки по вертикали указываются женские гаметы, по горизонтали — мужские. В квадраты решетки вписываются образующиеся сочетания гамет. Эти сочетания соответствуют генотипам зигот. Решетка Пеннета особенно удобна при анализе наследования признаков сложных гибридов.
Рассмотрение приведенной схемы показывает, что расщепление по фенотипу 3 : 1 и генотипу 1:2:1 может быть осуществлено лишь при определенных условиях. Во-первых, должно происходить равновероятное образование в мейозе обоих типов гамет т. е. гамет, несущих аллель А, и гамет с аллелью а. Во-вторых, должна иметь место равновероятная встреча и сочетание этих гамет при оплодотворении. Оба эти условия, как мы знаем из предыдущей главы, обеспечиваются с большей точностью при большем числе наблюдений. В первом делении мейоза осуществляется редукция числа хромосом с расхождением гомологичных партнеров к полюсам, причем вероятность отхождения хромосом к тому или другому полюсу одинакова, как и вероятность созревания и развития гамет всех сортов. При большом числе гамет обеспечивается также и равновероятная встреча их при оплодотворении. К этим условиям необходимо добавить и еще одно — равную выживаемость всех типов зигот и развивающихся из них особей.
Анализируя моногибридное скрещивание, мы не обращали внимания на то, какое из растений было материнским и какое — отцовским. Влияет ли на свойства гибрида и на характер расщепления в его потомстве то, что материнское растение будет нести доминантный признак, а мужское — рецессивный, и наоборот? Еще до Менделя гибридизаторы заметили, что направление скрещивания обычно не влияет на признаки гибрида. Это давало основание предполагать равное участие женского и мужского полов в передаче наследственных факторов. Мендель подтвердил эти наблюдения. Доминантный признак проявляется у гибрида независимо от того, привносит ли этот признак материнское или отцовское растение. Для признаков гороха, наследование которых изучал Мендель, указанное положение было правильным.
Но надо сказать, что иногда имеются различия в передаче наследственных свойств со стороны материнского или отцовского организма, с этим явлением мы познакомимся позднее. Поэтому направление скрещивания все же принято указывать. Скрещивания двух форм между собой в двух разных направлениях называют реципрокными. Так, при скрещивании двух форм Р1 и Р2 в одном направлении Р1 выступает как материнская форма, Р2 — как отцовская (P1XP2), а во втором — Р2 как материнская, Р1 — отцовская (P2XP1).