Анализ наследования одной аллельной пары в моногибридном скрещивании позволил Менделю и его последователям понять наследование двух и более пар признаков при дигибридном и полигибридном скрещиваниях.
Из всего ранее изложенного по этому поводу можно сделать следующие выводы:
- альтернативные признаки определяются аллелями гена, находящимися в гомологичных хромосомах;
- в каждой паре аллелей одна может быть доминантной, другая — рецессивной;
- во втором поколении моногибридного скрещивания происходит расщепление по доминантным и рецессивным признакам в определенном отношении; при полном доминировании расщепление по фенотипу 3:1, при неполном — 1:2:1, последнее отношение соответствует расщеплению по генотипу. Расщепление в F2 открыло принципиально важное явление, а именно: признаки скрещивающихся форм не пропадают в поколениях, а сохраняются;
- при дигибридном скрещивании обе аллельные пары, находящиеся в негомологичных парах хромосом, наследуются независимо друг от друга и вступают во все возможные комбинации с равной вероятностью, давая расщепление во втором поколении по фенотипу 9:3:3: 1 и по генотипу 1:2:2:4:1:2:1:2:1;
- явление расщепления и независимой комбинации разных пар аллелей генов обусловлено тем, что эти гены находятся в негомологичных хромосомах, которые случайно комбинируются при расхождении к полюсам в анафазе I мейоза.
Эти основные положения, установленные генетическим и цитологическим методами, позволяют сделать следующий шаг в анализе закономерностей наследования: выяснить их общее значение при полигибридном скрещивании.
Рассмотрим, например, тригибридное скрещивание. Можно заранее сказать, что основные закономерности наследования, установленные при моногибридном и дигибридном скрещиваниях, являются общими и для наследования при тригибридном скрещивании. Мендель для такого рода скрещивания избрал следующие три пары альтернативных признаков семян: гладкая — морщинистая форма семян, желтая — зеленая окраска семядолей, а также серо-коричневая и неокрашенная кожура, причем первыми в парах названы доминантные признаки, а вторыми — рецессивные.
В этом скрещивании материнское растение было гомозиготным по всем трем доминантным признакам, т. е. имело генотип ААВВСС, а отцовское — гомозиготно по трем рецессивным aabbcc. Все три гена находятся в трех негомологичных хромосомах. Тригибрид первого поколения имел структуру A/a B/b C/c. По внешнему виду семена тригибрида полностью походили на материнские, т. е. имели все три доминантных признака: гладкую форму, желтую окраску семядолей и серо-коричневую окраску семенной кожуры.
При указанной степени гетерозиготности гибриды F1 образуют уже восемь сортов гамет как женских, так и мужских, а именно: ABC, AbC, АВс, аВС, Abe, аВс, аbС и abc. При оплодотворении в результате сочетания восьми сортов женских и восьми сортов мужских гамет во втором поколении образуется 64 комбинации. В этом можно легко убедиться, воспользовавшись либо решеткой Пеннета, либо математическим способом перемножения вероятностей, как это мы делали раньше при дигибридном скрещивании. В результате подсчета обнаруживается, что у тригибрида расщепление в F2 по фенотипу включает 8 классов в определенном числовом отношении: 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1.
Очевидно, для того чтобы выявить все 8 классов расщепления по фенотипу, необходимо иметь достаточно большое число растений в F2. И Мендель отмечает, что из всех опытов опыт с тригибридом потребовал наибольшего количества времени и труда. Чтобы убедиться в том, что тригибрид действительно образует 8 типов гамет в равных количествах, можно воспользоваться известным нам методом анализирующего скрещивания, который не требует столь большого числа потомков, как это необходимо для обнаружения восьми фенотипических классов в F2. Скрещивая тригибрид с формой, гомозиготной по всем трем рецессивным признакам (A/a B/b C/c x a/a b/b c/c), мы получим расщепление в отношении 1 : 1 : 1 : 1 : 1 : 1 : 1 : 1.
Если рассчитать расщепление в F2 по фенотипу отдельно для каждой пары альтернативных признаков, то расщепление в каждом случае окажется равным 3:1. Это исходное отношение, как мы знаем, обеспечивается точным цитологическим механизмом независимого расхождения пар хромосом в мейозе. Именно это числовое отношение в расщеплении по каждой паре признаков позволило Менделю и в тригибридном скрещивании подтвердить принцип независимого сочетания признаков в потомстве гибрида.
Принцип независимого поведения разных пар альтернативных признаков в расщеплении по фенотипу в F2 при полном доминировании может быть выражен формулой (3 + 1)n, где n — число пар альтернативных признаков.
Исходя из этой формулы, можно высчитать число ожидаемых классов в расщеплении по фенотипу при любом числе пар признаков, взятых в скрещивание:
- моногибридное скрещивание (3+1)1 = 3 : 1, т. е. 2 класса,
- дигибридное скрещивание (3+1)2 = 9: 3 : 3 : 1, т. е. 4 класса,
- тригибридное скрещивание (3 + 1)3 = 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1, т. е. 8 классов и т. д.
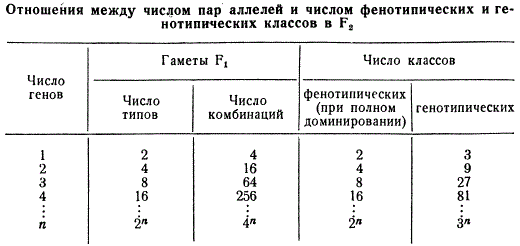
Иначе, говоря, число фенотипических классов при расщеплении в F2 может быть выражено формулой 2n, где основание 2 указывает на парность генов (аллельность), находящихся в одной паре гомологичных хромосом, а степень n — число аллельных пар в негомологичных хромосомах, по которым различаются взятые в скрещивание родительские формы. Поэтому при моногибридном скрещивании число классов расщепления по фенотипу равно 21 = 2, при дигибридном 22 = 4, тригибридном 23 = 8, тетрагибридном 24 = 16 и т. д.
Таким же образом можно рассчитать число типов гамет, образующихся у любого полигибрида первого поколения (F1), и число комбинаций гамет, дающих различные генотипы в F2.
- у моногибрида A/a образуются два сорта гамет, или 21
- у дигибрида A/a B/b — четыре, или 22
- у тригибрида A/a B/b C/c — восемь сортов гамет, или 23
Следовательно, число различных типов гамет, образуемых гибридом F1, также может быть выражено формулой 2n, где n — число пар аллельных генов, по которым различаются скрещиваемые формы.
Так как при моногибридном скрещивании у гибрида F1 образуются два сорта как женских, так и мужских гамет, то очевидно, что при этом возможно образование 4 комбинаций: 1А/А : 2А/a : 1a/a, т.е. 41.
При дигибридном скрещивании таких сочетаний будет 42 = 16, при тригибридном 43 = 64 и т. д., т. е. число возможных комбинаций гамет выражается формулой 4n, где основание 4 отражает число возможных комбинаций мужских и женских гамет в моногибридном скрещивании, а n — число пар аллелей.
Число генотипических классов рассчитывают при любом числе аллельных пар, по которым различаются родительские формы, взятые в скрещивание. В потомстве моногибрида расщепление по генотипу включает три класса: 1А/А : 2А/a : 1a/a.
При дигибридном скрещивании в F2 число генотипов соответствует 9 классам:
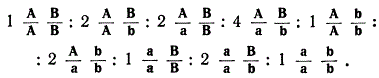
При тригибридном скрещивании число генотипических классов будет равно 33.
Итак, число генотипических классов можно определить по формуле 3n, где n — число аллельных пар. Таким образом, зная число пар аллелей при полигибридном скрещивании, можно рассчитать число типов гамет, образующихся у гибрида F1 число их сочетаний при оплодотворении, а также число фенотипических и генотипических классов.
Следует еще раз подчеркнуть, что все эти расчеты правомочны при том важном условии, если гены находятся в негомологичных хромосомах. Однако известно, что число последних для каждого Вида организмов является относительно небольшим и постоянным. Так, у гороха имеется всего 7 пар гомологичных хромосом, у человека — 23, у мушки дрозофилы — 4 и т. д. Следовательно, возможно одновременное независимое наследование лишь стольких генов, сколько пар гомологичных хромосом имеется у организмов Данного вида. Поскольку у гороха имеется 7 пар хромосом (n = 7), то у гибрида возможно независимое сочетание не более семи одновременно взятых в скрещивание пар признаков. При этом в потомстве гибридного растения гороха в F2 при полном доминировании возможно появление 27 различных фенотипических классов и 37 генотипических классов. Общее число возможных сочетаний гамет составит 47. У классического для генетических исследований объекта — дрозофилы по независимо наследующимся парам признаков возможно только тетрагибридное скрещивание.
Очевидно, что если в скрещивание будет взято число аллельных пар, большее, чем имеющееся у данного организма число пар гомологичных хромосом, то третий закон Менделя не осуществится. В этом случае вступят в действие другие закономерности.
На первый взгляд это ограничение закона независимого сочетания генов может создать впечатление ограниченности наследственной изменчивости в силу небольшого числа возможных комбинаций гамет. Рассмотрим для примера возможный размах комбинативной изменчивости, возникающей в силу свободной комбинации гамет у человека. Допустим, что в каждой из 23 пар хромосом человека имеется только по одной паре аллелей: A/a, B/b, C/c, D/d, E/e и т.д.
При таком расчете число различных сортов гамет выразится величиной 223, число классов расщепления по фенотипу — 223, по генотипу — 323, а число возможных комбинаций гамет — 423, или 246.
Для иллюстрации приводится число типов гамет и их сочетаний у индивидуумов, гетерозиготных по разному числу генов, от 1 до 23.
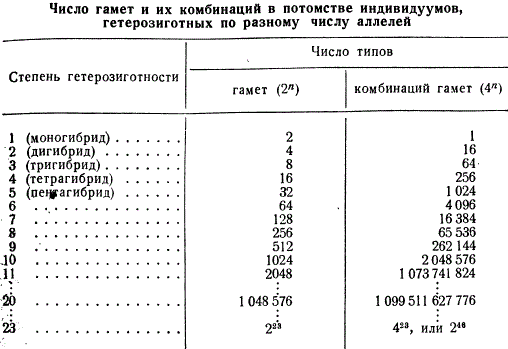
Подсчеты показывают, насколько велико число комбинаций гамет, которое может обеспечить наследственную изменчивость у человека, если различие одновременно касается 23 пар генов. Надо заметить, что в одном эякуляте мужчины число сперматозоидов колеблется от 2х109 до 5×109. Следовательно, даже при таком огромном количестве сперматозоидов в одном эякуляте далеко не все указанные выше типы гамет могут встретиться.
Эти иллюстративные вычисления отражают лишь незначительную долю той изменчивости, которая в действительности возможна при свободном сочетании гамет. Во-первых, в каждой паре гомологичных хромосом может быть более одного гена. Во-вторых, при рассмотрении расщепления мы исходили из того, что разные пары аллелей в своем определении признаков не взаимодействуют между собой; на самом же деле это не так: в процессе индивидуального развития организма гены взаимодействуют. Существует и ряд других причин, которые могут приводить к увеличению изменчивости генотипов и фенотипов.
Несмотря на указанные моменты и тот факт, что каждый организм получает половину хромосом от матери, а половину от отца, наследственное сходство между родителями и потомством надежно обеспечивается. Комбинативная изменчивость представляет собой перекомбинацию наследственных возможностей родителей. На ограниченном числе потомков она не может быть выявлена. Лишь на большом числе потомков комбинативная изменчивость реализуется с большой полнотой.
В заключение изложения классической схемы анализа наследования признаков и свойств организмов следует сказать о том, в какой мере этот анализ можно и следует применять. Закономерности наследования, установленные Менделем на горохе, являются общими для всех организмов, имеющих половой процесс. Однако проявление их может видоизменяться в зависимости от объекта и влияния факторов внешней среды. Каждое закономерное явление природы осуществляется при определенных условиях. Если эти условия не способствуют его проявлению, то оно или совсем не проявится, или проявление его изменится. Но если сам метод обнаружения закономерного явления объективен, то с его помощью можно вскрыть новый характер проявления закономерности при измененных условиях. В этом сила научного метода.