В селекции животных и растений особое место занимает явление гибридной мощности, или гетерозиса, которое заключается в следующем.
При скрещивании разных рас, пород животных и сортов растений, а также инбредных линий гибриды F1 по ряду признаков и свойств часто превосходят исходные родительские организмы. Скрещивание гибридов между собой ведет к затуханию этого эффекта в следующих поколениях. В настоящее время гетерозис установлен для всех изученных видов.
Хотя эффект гетерозиса известен с древнейших времен, его природа до сих пор остается мало изученной. Первое толкование биологического значения гетерозиса в эволюции животных и растений и попытка объяснить механизм этого явления принадлежали Ч. Дарвину. Он систематизировал большое количество фактов и сам ставил опыты по изучению гетерозиса. По мнению Ч. Дарвина, гетерозис служит одной из причин биологической полезности скрещивания в эволюции видов. Перекрестное оплодотворение поддерживается естественным отбором именно потому, что оно служит механизмом сохранения наибольшей гетерозисности,
Глубокий научный анализ явления гетерозиса стал возможен только с начала XX в. после открытия основных генетических закономерностей. При рассмотрении гетерозиса мы будем касаться возникновения его при скрещивании преимущественно инбредированных линий одного вида, поскольку в этом случае легче выяснить его генетический механизм.
С начала нашего столетия на кукурузе стали проводить систематическое исследование скрещиваний между инбредными линиями. При этом Г. Шеллом было показано, что скрещивание некоторых линий дает гибридные растения, более урожайные по зерну и вегетативной массе, чем исходные линии и сорта. Приведены опытные данные, показывающие низкую урожайность инбредных линий, значительное повышение урожайности в F1 и снижение в F2 при самоопылении растений F1.
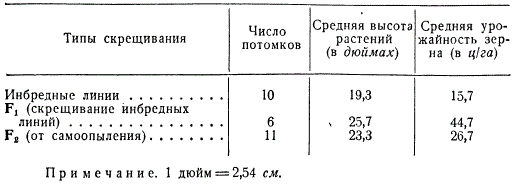
Показатели мощности растений при разных типах скрещивания у кукурузы
За последние 30 лет в ряде стран, в том числе и в нашей, посев гибридными семенами стал основным приемом производства кукурузы как на зерно, так и для силоса. Для получения гибридных семян сначала создают инбредные линии из лучших сортов, отвечающих требованиям данного климатического района (инбредная линия создается в течение 5—6 лет путем самоопыления). При отборе линий оценивают их качества и свойства, которые необходимо получить у будущего гибридного организма. Инбридинг в линиях не может быть эффективным, если он не сопровождается отбором.
Создав большое число инбредных линий, приступают к скрещиванию между ними. Межлинейные гибриды первого поколения оценивают по эффекту гетерозиса; исходя из этого показателя отбирают линии с лучшей комбинационной ценностью и затем размножают их в больших масштабах для производства гибридных семян. Работа по созданию инбредных линий и оценке их комбинационной ценности постоянно ведется в селекционных учреждениях. Чем больше создано ценных линий, тем вернее й скорее можно отыскать лучшие гибридные комбинации с необходимым сочетанием свойств.
При получении гибридных семян для производственных целей исходные линии, дающие при скрещивании наибольший эффект гетерозиса, высевают рядами, чередуя материнские и отцовские формы. Чтобы обеспечить опыление между ними, с материнских растений удаляют мужские соцветия (метелки). Теперь разработана новая схема производства гибридных семян с использованием цитоплазматической мужской стерильности, что позволило значительно сократить труд на удаление метелок у растений материнской линии. Таким путем получают простые межлинейные гибриды кукурузы. Этот метод в принципе является общим для семеноводства гибридов различных перекрестноопыляющихся растений.
В настоящее время в практике сельского хозяйства простые межлинейные гибриды кукурузы не используются, так как затраты на получение таких семян не окупаются. Теперь широко внедряется в практику посев семян двойных межлинейных гибридов. Последние получают путем скрещивания двух простых гибридов, проявляющих гетерозис.
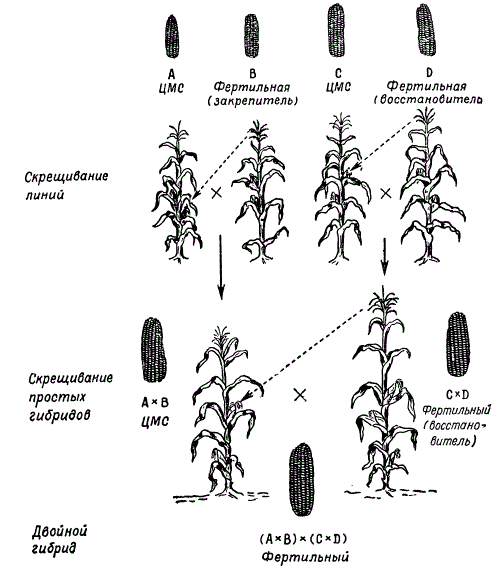
Схема получения двойных гибридов кукурузы с использованием цитоплазматической мужской стерильности
По данным М. И. Хаджинова и Г. С. Галеева, результаты оценки продуктивности разных гибридов — межсортовых, сортолинейных и двойных межлинейных — показывают, что наиболее продуктивны двойные межлинейные гибриды.
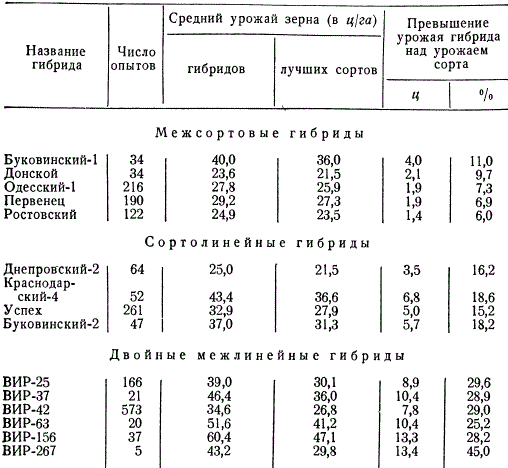
Продуктивность гибридов кукурузы в сравнении с сортами
Подбор простых гибридов для получения наиболее продуктивных двойных гибридов является важным этапом их селекции. Лучшие результаты дает скрещивание линий, происходящих из различных сортов. Так, например, если один простой гибрид получен от скрещивания инбредных линий двух сортов А X В, а другой — от скрещивания линий других сортов С х D, то двойной гибрид (А X В) х (С X D) дает гетерозис чаще, чем если бы двойной гибрид был получен от скрещивания простых гибридов, происходящих от линий одного сорта: (А х А1 х (А2 х А3) или (В х B1) х (В2 х В3).
Для успешного развития гибридного семеноводства необходимо исследовать, во-первых, как долго следует вести инбридинг, чтобы получить гомозиготные линии, и, во-вторых, разработать методы более быстрой оценки их комбинационной ценности.
Все, что было сказано в отношении кукурузы, по-видимому, является общим для получения гибридов у других высших перекрестноопыляющихся растений, а также у животных. В настоящее время в птицеводстве и в свиноводстве ряда стран широко используется скрещивание инбредных линий, происходящих из одной или разных пород. Следует особо подчеркнуть, что широкое использование гибридов в животноводстве возможно только при высоком уровне племенной работы и наличии ценных пород, то совершенно ясно, что в большинстве случаев инбредные линии в среднем будут иметь всегда более низкие показатели, чем сорта. О наличии гетерозиса следует говорить лишь в том случае, когда межлинейный гибрид превосходит не только родителей (линии), но и сорта, или породы, от которых произошли эти линии.
Как мы знаем, гены детерминируют свойства организма на всех стадиях онтогенеза начиная с момента оплодотворения. Гены ядра ооцита способны детерминировать свойства цитоплазмы яйцеклетки еще до оплодотворения. Характер реализации генотипа изменяется в зависимости от свойств цитоплазмы зиготы. Таким образом, проявление гетерозиса у гибрида зависит также и от свойств цитоплазмы. Роль цитоплазмы в определении гетерозиса доказывается следующим образом. При реципрокном скрещивании двух линий А X В и В х А гетерозис по одним и тем же свойствам нередко проявляется у гибридов только одного из скрещиваний и не проявляется у гибридов другого.
Проявление гетерозиса обусловлено индивидуальным развитием гибрида. В онтогенезе он реализуется неравномерно. На одних стадиях онтогенеза проявляется гетерозис по одним признакам, а на других — по другим или только по некоторым. Так, в раннем возрасте у одного и того же гибрида может наблюдаться гетерозис в отношении скорости роста отдельных частей организма и повышенной устойчивости к заболеваниям, но его может не быть, например, в отношении устойчивости к неблагоприятной температуре. Гетерозис по этому свойству может проявляться позднее.
Сильное влияние на проявление гетерозиса оказывают также факторы среды, в которых развивается гибридный организм. Есть основание считать, что гетерозис по жизнеспособности и устойчивости к заболеваниям лучше выявляется при неблагоприятных для развития условиях среды. Еще Ч. Дарвином было высказано предположение, что гетерозис у гибридов обусловлен более широкой нормой приспособительных реакций. Исследования последних лет показали, что гибриды F1 в сравнении с инбредными линиями характеризуются большим разнообразием процессов обмена, участием большего числа различных метаболитов, ростовых веществ и энзимов. Гетерозис проявляется не только в целом организме, но и на клеточном уровне.
Выяснение генетических механизмов гетерозиса остается нерешенной проблемой. В настоящее время имеются три гипотезы, пытающиеся объяснить возникновение гетерозиса:
- гетерозиготное состояние по многим генам,
- взаимодействие доминантных благоприятных генов,
- сверхдоминирование — гетерозигота превосходит гомозиготы.
Как мы уже говорили, при скрещивании гомозиготных инбридированных линий гибриды первого поколения по многим генам приобретают гетерозиготное состояние. При этом действие вредных рецессивных мутантных аллелей подавляется доминантными аллелями обоих родителей. Схематически это можно представить так: одна инбредная линия в гомозиготном состоянии имеет рецессивную аллель одного гена ааВВ, а вторая — другого гена ААbb. Каждая из этих рецессивных аллелей генов в гомозиготном состоянии определяет какую-нибудь недостаточность, которая снижает жизнеспособность инбредной линии. При скрещивании линий ааВВ X ААbb у гибрида объединяются доминантные аллели обоих генов (АаВb). Гибриды F1 проявят при этом по указанным генам не только гетерозис, но и единообразие. В F2 число особей с двумя доминантными генами в гетерозиготном состоянии будет лишь 4/16, поэтому гетерозисными оказываются не все особи. В дальнейших поколениях число гетерозигот сокращается, а число гомозигот увеличивается. В силу этих причин гетерозис в последующих поколениях затухает. Такова схема одной из генетических гипотез гетерозиса.
Мы привели пример с двумя генами, но физиологические свойства определяются огромным числом генов. Кроме того, неверно считать, что доминантные аллели всегда положительно, а рецессивные отрицательно влияют на жизнеспособность организма и его продуктивность. Доминантные аллели дикого типа чаще оказывают благоприятное действие, чем рецессивные. Это можно объяснить тем, что неблагоприятные доминантные мутации подвергаются более жесткому отбору — они уже в зиготе и на эмбриональных стадиях подвергаются элиминации, и отбором сохраняются лишь благоприятные. И так как само свойство доминирования гена эволюционирует под контролем отбора, то аллели дикого типа и оказываются более благоприятными для организма. Поэтому подбор в гибридной комбинации доминантных аллелей может скорее обеспечить гетерозис. Если бы гетерозис обусловливался простым набором доминантных аллелей, имеющихся в популяции, то этот набор было бы легко составить путем ряда скрещиваний и получить гетерозисные сочетания. Весьма возможно, что часть пород и сортов выводилась именно путем скрещивания и комбинаций линий, имеющих набор благоприятных доминантных аллелей. Но до сих пор не удавалось закрепить гетерозис в гибридной комбинации F1, т. е. получить не расщепляющиеся в F2 формы.
Д. Джонсом еще в 1917 г. было предложено дополнение к изложенной гипотезе. Согласно Д. Джонсу, разные гены, дающие в сочетании гетерозис, находятся в пределах одной группы сцеплений: например, AbcdE — в одной, а соответственно aBCDe — другой гомологичной хромосоме. Именно это обстоятельство, с точки зрения Д. Джонса, затрудняет отбор в F2 полностью гомозиготных форм по благоприятным доминантным генам, дающим сочетании гетерозис. Для осуществления такого сочетания доминантных аллелей необходимо, чтобы в этой паре хромосом AbcdE//aBCDe произошел минимум двойной кроссинговер, который привел бы к возникновению хромосомы с одними доминантными аллелями ABCDE. Малая вероятность одновременного осуществления такого события в нескольких хромосомах и является, по мнению Д. Джонса, причиной, затрудняющей закрепление гетерозиса. Гибрид же F1 содержит все эти доминантные аллели, и поэтому у него наблюдается гетерозис.
Такова схема объяснения гетерозиса согласно второй гипотезе — гипотезе взаимодействия доминантных благоприятных генов, которую называют иногда также гипотезой набора кумулятивных доминантных аллелей, а сам гетерозис при этом — мутационным. Эта гипотеза в целом исходит из представления о простом суммировании эффекта доминантных аллелей с комплементарным действием.
Третья гипотеза исходит из того, что гетерозиготное состояние аллелей имеет превосходство над гомозиготным (АА < Aa > аа). Здесь можно допустить благоприятное трансположение аллелей дикого типа и мутантных аллелей, которое каким-то образом усиливает эффект действия генов. Указанное объяснение гетерозиса называют гипотезой сверхдоминирования.
Ни одна из трех гипотез не может считаться единственно правильной. Сейчас преждевременно отдавать предпочтение той или другой. Вероятно, они все окажутся правильными, но для разных случаев. Наверное, каждый из механизмов, предусматриваемых этими гипотезами, играет роль в определении гибридной мощности. Гетерозис является сложным явлением как по механизму возникновения, так и по проявлению его в онтогенезе.
Очевидно, окончательный вывод о генетических механизмах гетерозиса можно будет сделать лишь после того, как раскроется картина взаимодействия генов в генетической системе на биохимическом и молекулярном уровнях. Как мы уже знаем, подавление проявления мутаций с помощью супрессоров позволяет исправлять нарушения в действии гена и его аллелей. Возможно, что явление гетерозиса есть суммарный результат работы не основных генов, определяющих развитие признаков организма, а набора супрессирующих генов в генотипе. Важным моментом изучения гетерозиса становится исследование плазменных отношений.
Основной задачей использования гетерозиса в селекции является закрепление его, т. е. сохранение эффекта гетерозиса в процессе воспроизведения гибрида. Решение этой задачи мыслится в нескольких аспектах: во-первых, закрепление гетерозиса путем перевода гибридного организма с полового размножения на апомиктическое, что, по-видимому, возможно для некоторых растений; во-вторых, закрепление гетерозиса путем перевода диплоидного гибрида, проявляющего гетерозис, в полиплоидное состояние. В этом случае гетерозиготная комбинация генов будет сохраняться дольше.
У вегетативно размножающихся растений поддержание ценных гибридных комбинаций, полученных половым путем, осуществляется вегетативным размножением (черенками, прививками, клубнями и т. д.).
Существует ряд других путей поддержания гетерозиса в ряду поколений, однако все они еще недостаточно разработаны и проверена.