Из оплодотворенной яйцеклетки, или зиготы, путем последовательных делений развивается зародыш, из оплодотворенной центральной клетки или клетки зачатка эндосперма развивается эндосперм.
После слияния спермия с яйцеклеткой, т. е. мужской и женской гаплоидных гамет, в оплодотворенной яйцеклетке восстанавливается диплоидное число хромосом. Первичное ядро эндосперма, являясь обычно продуктом слияния трех ядер — двух полярных и одного спермия — имеет триплоидное число хромосом. В редких случаях, когда эндосперм образуется без оплодотворения, его клетки имеют диплоидное число хромосом, а иногда и гаплоидное (представители семейства Onagraceae). Наконец, у тех растений, у которых при образовании вторичного ядра сливаются не два, а несколько ядер (3, 4, 7, 8 и даже 14), эндосперм может быть более или менее полиплоидным. После оплодотворения яйцеклетка и центральная клетка зародышевого мешка становятся способными к активному развитию. Двойное оплодотворение отсутствует у всех архегониальных растений и свойственно лишь покрытосеменным.
Во время развития зародыша и эндосперма зародышевый мешок нуждается в особенно обильном притоке питательных веществ, который осуществляется за счет извлечения их из окружающих материнских тканей, а также благодаря непосредственному потреблению зародышевым мешком тканей нуцеллуса и покровов семяпочки. Этому способствуют различного типа гаустории (присоски), развитые у многих растений. Под воздействием интенсивного обмена веществ размеры зародышевого мешка значительно увеличиваются. Наряду с этим стимулируется дальнейший рост и развитие семяпочки.
Период времени, протекающий от оплодотворения до образования зародыша и эндосперма, у разных растений неодинаков, колеблясь от 10—15 дней (у кок-сагыза, салата и др.) до нескольких недель (у злаков, хлопчатника и др.) и даже до нескольких месяцев (у черного саксаула и др.).
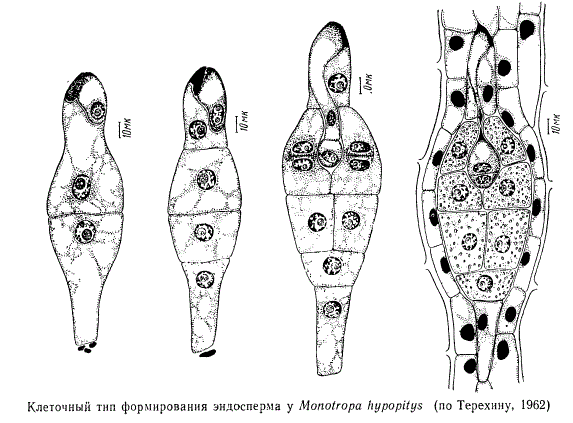
После двойного оплодотворения чаще всего первым делится первичное ядро эндосперма (продукт слияния вторичного — центрального ядра зародышевого мешка со спермием). Эндосперм у разных растений формируется неодинаково. Различают три основных типа развития эндосперма: ядерный, или нуклеарный, клеточный, или целлюлярный, и базальный, или тип Helobiae. В каждом из этих типов имеется ряд более или менее значительных модификаций.
Ядерный тип образования эндосперма характеризуется тем, что вслед за делением первичного ядра эндосперма не происходит немедленного образования клеточных перегородок, а возникает большее или меньшее число ядер, свободно лежащих в общей цитоплазме.
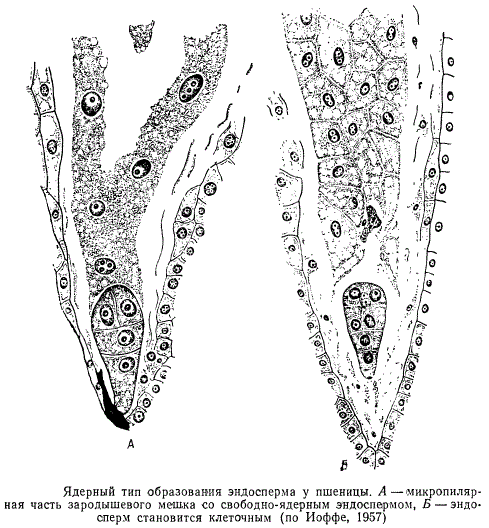
Формирование клеточных перегородок при ядерном типе развития эндосперма наступает позднее, начинаясь либо в микропилярной части зародышевого мешка, либо по всей периферии его. В редких случаях при ядерном типе эндосперма клеточные оболочки вообще не закладываются. Клеточный тип, напротив, характеризуется тем, что после первого и каждого из последующих делений ядер эндосперма немедленно появляются клеточные перегородки.
Базальный тип имеет промежуточный характер между ядерным и клеточным. Он отличается тем, что возникающая сразу после первого деления ядра эндосперма клеточная перегородка делит центральную клетку на две неравные части: большую верхнюю, или микропилярную, и маленькую нижнюю, или базальную. В микропилярной части эндосперма сначала образуется ряд свободных ядер и только позднее между ядрами появляются клеточные перегородки; в нижней же части либо образуются две клетки, либо небольшое число свободных ядер, либо делений вообще больше не происходит. Эта антиподальная часть эндосперма, или его базальный аппарат, функционирует часто как гаусторий.
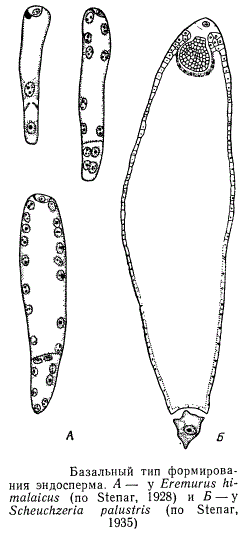
Эндоспермальные гаустории формируются также при клеточном, а иногда и при ядерном типе развития эндосперма, возникая как в микропилярной, так и в халазальной и даже в латеральной части зародышевого мешка.

Гаусториями называются образования, состоящие из одно- или многоядерных клеток с густым содержимым и часто с гипертрофированными клеточными ядрами. В одних случаях гаустории состоят из нескольких более или менее разросшихся и разветвленных клеток и даже целой ткани, в других — только из одной клетки, сильно разрастающейся и дающей боковые отростки.
Наряду с эндоспермальными гаусториями в семяпочках покрытосеменных известны и другие типы гаусторий, образованных самим зародышевым мешком, антиподами, синергидами, подвеском зародыша. Гаустории весьма сильно варьируют по своему облику и строению, но роль их, по-видимому, одна и та же; они приспособлены к выполнению питательной и секреторной функции. Проникая своими отростками и боковыми ветвями глубоко в ткани семяпочки, плаценты и даже завязи, гаустории извлекают из материнского растения питательные вещества для роста и формирования зародыша. Эндоспермальные гаустории, как и гаустории любого другого типа, способствуют повышению интенсивности обмена веществ во время роста и развития зародыша, так как облегчают приток к нему питательных веществ из окружающих тканей материнского растения.
Примером гаусторий антиподального происхождения могут служить гаустории злаков. Комплекс антипод образует своеобразное подобие ткани, внедряющейся в нуцеллус. Клетки нуцеллуса, непосредственно примыкающие к клеткам антиподального комплекса, разрушаются.
При гибридизации и самоопылении перекрестников, а также под воздействием неблагоприятных внешних условий развитие эндосперма может преждевременно приостанавливаться или с самого начала происходит ненормально. Это, в свою очередь, сразу сказывается на формировании зародыша и приводит к образованию щуплых семян с недоразвитым зародышем.
В сформированном виде эндосперм зрелого семени (тех растений, у которых он сохраняется) представлен тканью, состоящей из более или менее крупных клеток, заполненных различными питательными веществами — крахмалом, маслом, сахарами, белком и т. д. Кроме того, в клетках эндосперма могут присутствовать дубильные вещества, алкалоиды, кристаллы оксалата кальция и т. д. Оболочки эндоспермальных клеток обычно тонкие, целлюлозные. У некоторых растений, у которых основное запасное вещество откладывается в оболочках, последние значительно утолщаются.
По своей консистенции эндосперм у одних растений более или менее жидкий или студенистый, у других — в той или иной степени твердый или даже роговой. Эндосперм бывает либо бесцветным, либо различно пигментированным.
В дальнейшем эндосперм претерпевает различные превращения. У одних растений он полностью ассимилируется зародышем в процессе развития последнего и в зрелом семени или отсутствует полностью, или представлен одним-двумя слоями клеток. К таким растениям принадлежат бобовые, тыквенные, сложноцветные и другие. В этих случаях запасные питательные вещества, необходимые для прорастания зародыша, накапливаются в клетках семядолей самого зародыша, и последний заполняет собой почти весь объем семени.
У других растений эндосперм сохраняется до созревания семени и потребляется зародышем лишь при его прорастании (злаковые, ситниковые, магнолиевые и др.). В этих случаях зародыши малы по объему, а основную часть семени занимает эндосперм. Между этими крайними примерами существует, конечно, много переходных форм, среди которых встречаются и такие, у которых объемы эндосперма и зародыша в зрелом семени одинаковые.
У некоторых растений (например, из семейства гвоздичных, перечноцветных) запасные вещества откладываются в нуцеллусе. Подобного рода запасная ткань называется периспермом.
Оплодотворенная яйцеклетка, или зигота, может делиться одновременно с делением первичного ядра эндосперма, но, как правило, зигота делится несколько позже (иногда значительно позже) — лишь после образования нескольких, а в некоторых случаях большого числа ядер или клеток эндосперма. Во время созревания зиготы она сама и ее ядро значительно увеличиваются в размерах, вакуоля постепенно исчезает, и вся клетка заполняется более или менее густым содержимым, концентрирующимся главным образом в апикальной ее части. По окончании периода созревания зигота начинает делиться, давая начало зародышу.
Имеется несколько классификаций типов развития зародыша, в основе которых лежат следующие признаки: способ заложения первой перегородки, делящей зиготу на две клетки, направления перегородок при последующих делениях, а также участие тех или иных клеток начинающего развиваться зародыша в формировании его органов.
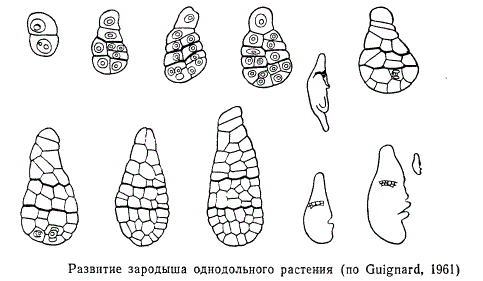
При развитии зиготы покрытосеменного растения возникает клеточное тело, которое обычно уже на начальных фазах своего развития расчленено на базальную часть — подвесок, или суспензор, и терминальную — тело зародыша. Независимо от способа заложения клеточных перегородок зародыши покрытосеменных растений на самых ранних стадиях внешне довольно единообразны и являются радиально симметричными. В этот период нет различий даже между зародышами двудольных и однодольных растений, эти различия появляются позже. У двудольных растений тело зародыша постепенно принимает форму шара, который в дальнейшем несколько уплощается, после чего происходит дифференциация на семядоли и осевую часть. Через такой зародыш можно теперь провести только две плоскости симметрии.

При развитии зародыша однодольных растений шарообразная стадия отсутствует, а зародыш из радиально симметричного становится билатерально симметричным, т. е. через него можно провести лишь одну плоскость симметрии. В некоторых случаях, например у злаков, это объясняется тем, что здесь очень рано нарушается равномерность ритма деления клеток зародыша— на вентральной его стороне деления задерживаются, в то время как на противоположной дорзальной стороне и в апикальной части зародыша они протекают интенсивно. В результате зародыш изгибается, а это в свою очередь обусловливает иной по сравнению с двудольными характер заложения органов. Единственная семядоля закладывается терминально, а точка роста занимает боковое положение. В других случаях не исключены и другие пути, приводящие к становлению своеобразного зародыша однодольных.
Несколько особняком в отношении формирования зародыша среди покрытосеменных растений стоит семейство Леоновых. У его представителей деление зиготы и последующие деления производных ядер не сопровождаются заложением клеточных перегородок. В результате образуется предзародышевая ценоцитная структура, очень сходная с тем, что имеет место у голосеменных растений. Лишь позже между ядрами такого ценоцита закладываются перегородки и возникают меристематические очаги, дающие начало зародышам. Рост и дифференциация зародыша происходят в процессе активного обмена веществ, осуществляющегося между ним и окружающей его средой. Выше уже говорилось о том, что этому способствуют различного рода гаусториальные образования, формируемые различными структурами зародышевого мешка, в первую очередь эндоспермом. Такого рода гаусториальные функции свойственны также и самому зародышу, в частности его подвеску. При помощи подвеска зародыш продвигается в ткань эндосперма и получает наиболее благоприятные возможности для его использования; при помощи подвеска зародыш может получать питательные вещества и непосредственно из материнских тканей. Облегчая поступление к зародышу питательных веществ из окружающих его тканей семяпочки, подвесок способствует повышению интенсивности обмена веществ на ранних стадиях роста и развития зародыша.
Подвески зародышей бывают одноклеточными и многоклеточными, клетки в них могут быть одноядерными или многоядерными. В некоторых случаях подвесок сильно разрастается и разветвляется, образуя подвесочный гаусторий. Иногда же подвесок совершенно не развивается или развивается крайне слабо.