Примеры экологических рядов: 1) гигрогенного (борового) и 2) трофогенного замещения (боры — дубравы). На их примерах мы в кратких чертах иллюстрировали методику сравнительно-экологического изучения растительности, позволяющую установить ведущие факторы, которые определяют собой все главнейшие черты состава, строения и производительности леса.
Если не углубляться в даль веков и ограничиться лесоводственно приемлемым масштабом измерения времени, т. е. сроками, в пределах которых местообитания обычно не претерпевают коренных изменений (исключения — поймы, летучие пески, овраги и другие места современной геологической деятельности), то ведущими факторами, определяющими основное разнообразие состава и производительности естественных лесов, будут: свет, тепло, вода и минеральные питательные вещества почвы. Шкалы экологических свойств древесных пород, составленные лесоводами, прежде всего характеризовали отношение пород к этим факторам. Именно их количественные изменения и должны явиться стержнями (осями, ординатами) классификации типов леса, так как наиболее существенные качественные черты лесов вызваны различной количественной напряженностью экологических факторов.
В число наиболее важных для растения факторов простои неорганической среды входит пища. Казалось бы, сюда должна быть помещена углекислота воздуха как важнейшее питательное вещество для зеленых растений. Но мы уже условились, что ведущими факторами признаем лишь те, которые вызывают существенное разнообразие состава леса и связаны со значительной амплитудой количественной стороны этих факторов, а углекислота воздуха почти не обнаруживает соответствующих свойств. Такие важные для растения элементы среды, как почвенная кислотность, концентрация почвенного раствора, соотношение питательных элементов в почве, соотношение влажности воздуха и почвенной влаги, периодичность сезонного увлажнения, контрастность (континентальность) тепловых условий, периодичность и качественные стороны освещения могут быть предусмотрены при более детальной классификации лесных местообитаний, в пределах тех крупных единиц, которые выделяются на основе четырех указанных ведущих факторов.
Почвенная аэрация тесно связана с увлажнением почвы и представляет собой по отношению к последнему фактор функционально подчиненный. Это видно из того, что степень устойчивости растений к недостатку почвенной аэрации (ризоанаэробность) соответствует степени влаголюбив растений, причем исключения из этого правила (ксерофильная сосна на болоте, менее влаголюбивые нитрофилы, растущие в ольсах, и т. и.) связаны с пластичностью корневой системы, способной ограничиваться поверхностным почвенным слоем и другими приспособлениями, позволяющими невлаголюбивым растениям выносить эти условия без асфикции (без корневого задыхания). Следовательно, нет надобности предусматривать особую классификационную ординату для почвенной аэрации. Ее изменения, существенные для леса, даны в ординате влажности и, как мы это уже показали на примере борового ряда, могут быть измерены в единицах падения бонитета от точки оптимальной производительности до любой точки в сторону нарастающего избыточного увлажнения.
Микоризное питание многих древесных пород и установленная в последнее время физиологами по крайней мере частичная возможность питания зеленых растений за счет органических веществ как будто обязывают ввести в категорию основных классификационных факторов также и органическое вещество почвы. Однако мы пока еще не можем указать ни одной древесной породы, которая требовала бы органического вещества в качестве обязательного (незаменимого) элемента питания. Прочие факторы, не связанные с четырьмя ведущими (например, ветровая коррозия деревьев, характерная для некоторых ветроударных местообитаний; влияние характерных для некоторых горных местообитаний ожеледей, значительно снижающих производительность насаждений из-за постоянного обламывания вершин и ветвей; различия в количестве углекислоты воздуха и т. п.), могут быть предусмотрены как дополнительные варианты в пределах основных категорий типов леса, классифицируемых по четырем основным условиям жизни растений.
При первом подходе к экологическому изучению лесов тепло и свет (космические факторы) могут быть объединены в одну рубрику климатических факторов, а влажность и химические условия плодородия почвы — в рубрику эдафических. Оставляя в стороне климатическую влажность (осадки, влажность воздуха и т. п.), исходят из положения, что эдафическая (почвенная) влага имеет для растений исключительное значение как источник питания. Для леса это положение следует считать верным, но с теми оговорками, которые мы сделаем ниже. Климатические же изменения лесов мы вправе рассматривать в этом случае прежде всего, как тепловые (термогенные), считая, что изменения, вносимые географическим разнообразием световых условий, являются в основном параллельными разнообразию тепловых условий.
Как показал Е. В. Алексеев, в основу классификации лесов должны быть положены эдафические оси, чате всего обусловливающие местное пространственное разнообразие лесов. Вместе с тем эдафически однородные местообитания зачастую имеют в разных климатических областях близкий лесорастительный эффект (например, боры и в значительной мере субори однотипны на широких пространствах таежной зоны). Климатические факторы обнаруживают более ощутимое разнообразие в составе и производительности лесов лишь при рассмотрении их в широких территориальных масштабах, да и то в результате сложных отвлечений от эдафических черт в лесорастительном эффекте и от других моментов.
Было бы неправильным считать, что в процессе упомянутых сопоставлений и отвлечений леса и их местообитания делятся на части. Речь идет не только о частях, а и о ведущих факторах, само представление о которых предполагает не разделение или суммирование, а более сложную связь ведущего фактора со всеми остальными членами их комплекса. Здесь нет ничего общего с формально описательным разделением или объединением лесов. Эдафические и климатические оси (ординаты) леса и их местообитания классифицируют, т. е. сопоставляют и сравнивают между собой разные их стороны. При помощи классификации выясняются экологические взаимосвязи, устанавливается экологическое сходство и различие как между географически близкими, так и между отдаленными растительными сообществами.
Эдафическая сетка типов леса
Эдафическая сетка (классификационная система) лесов и местообитаний наглядно иллюстрирует те черты сходства и различия лесов, которые вызваны эдафическими особенностями местообитаний. Внешняя форма (сетка, диаграмма, шкала) служит только для того, чтобы наглядно показать единство двух классификационных ординат, лежащих в основе эдафического замещения: ординаты трофности (трофогенное замещение) и ординаты увлажнения (гигрогенное замещение) или, другими словами, показать единство сходства и различия лесов, вызванное количественным изменением химических условий плодородия (трофности) и увлажнения.
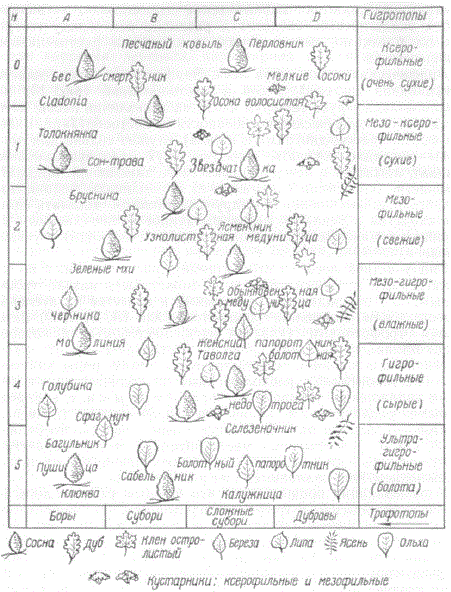
Эдафическая сетка (классификационная схема) типов леса. Нанесены условные знаки древесных пород и названия более распространенных представителей живого напочвенного покрова, характерных для Полесья и Лесостепи
Ряд участков леса (местообитаний), расположенных по количественным степеням трофности, может быть назван трофогенным, или порожденным различиями в качестве питательных веществ почвы. Отдельные члены этого ряда (А, В, С, D) называются трофотопами (участки леса, местообитания которых имеют одинаковое в своих пределах и отличающееся от других почвенное плодородие). Соответственно этому ряды участков леса (местообитаний), расположенные по количественным степеням увлажнения, называются гигрогенными, т. е. порожденными различиями в количестве эдафического (почвенногрунтового) увлажнения. Отдельные члены гигрогенного ряда — 0, 1, 2, 3, 4, 5 — участки леса, местообитания которых характеризуются одинаковым в своих пределах и отличающимся от других увлажнением. Они называются гигротопами.
Классификация предполагает, что каждый участок леса является одновременно и трофо- и гигротопом и по существу эти две классификационные единицы представляют собой лишь две стороны одного и того же местообитания — эдатопа. Эдатоп объединяет лесные участки одинаковых эдафических условий. Следовательно, упомянутую двумерную классификационную систему следует признать эдафической сеткой, или сеткой эдатопов. Отдельная классификационная единица — эдатоп, например, В2, представляет собой единство трофотопа В и гигротопа 2. Классификация дает меру почвенного (эдафического) плодородия местообитаний, исходя из их лесорастительного эффекта (индикаторов) как решающего критерия для оценки местообитаний в единстве двух упомянутых его сторон — почвенного плодородия и увлажнения.
Ряды увлажнения и трофности (почвенного плодородия) были рассмотрены нами. Если на эдафическую сетку нанести боровой ряд, он займет се первый вертикальный столбец А.
Рассмотренный нами ряд трофогенного замещения свежих по увлажнению местообитаний расположится во втором горизонтальном ряду эдафической сетки под цифрой 2. Все остальные ее места могут быть заполнены на основе фактического материала, а также путем интер- и экстраполяции.
В основе эдафической сетки лежат закономерные явления перехода количественных изменений в качественные и порождения новыми качествами новых же количественных изменений. Эти переходы были нами иллюстрированы на примере борового ряда и ряда трофогенного замещения. Единство организмов и среды позволяет рассматривать растительные сообщества и их местообитание неразрывно, как целостные явления. Нарастание количества влаги в гигрогенных рядах, как и нарастание количества минеральной пищи в рядах трофогенного замещения, вызывает многочисленные качественные скачки — переходы, отражающиеся в каждом ряду появлением и исчезновением видов растений в древесном и кустарниковом ярусах, в травяном, моховом и лишайниковом покровах. Кроме того, те же узловые линии меры, переходы количества в качество, свойственны также структуре и производительности древостоев. Эдафическая сетка как координатная система служит для иллюстрации влияния на растительность количественных градаций двух основных качеств эдафической среды — увлажнения и трофности. На эдафическую сетку могут быть нанесены показатели, отражающие качественные особенности состава растительности, а также и количественные особенности растений, виды напочвенного покрова (бонитеты древесных пород).
Лесные растения — экологические индикаторы
Иногда считают, что самым лучшим индикатором местообитания является биоценоз как целое и что отдельные признаки ценоза, отдельные растения в этом отношении уступают целому. Но в действительности наиболее глубокую и разностороннюю оценку местообитания мы можем получить, исходя из отдельных признаков ценоза, расчленяя его на отдельные части и рассматривая каждую часть в отдельности. Насаждение как целое, сплошь и рядом затушевывает для нас многие существенные стороны местообитания. Зачастую оно стирает внешние проявления важных сторон местообитания, оценка которых становится сложной и трудной.
Так, под сомкнутыми буковыми, пихтовыми и еловыми насаждениями из-за недостатка освещения живой напочвенный покров очень часто совсем отсутствует (например, мертвопокровные ценозы с господством бука, носящие латинское название Fagetum nudum, что означает «голый бучинник»). В грабняках на более бедных почвах такие великолепные индикаторы этих почв, как орляк, ракитник, дрок и т. и., являющиеся светолюбивыми спутниками сосны, исчезают. В одних случаях насаждение (фитоценоз) устраняет ценные признаки местообитания, в других, наоборот, может способствовать появлению таких признаков, по которым без глубокого понимания причин, обусловливающих их появление, можно составить ложное представление о существенных особенностях местообитания (например, появление зеленых мхов и грушанок на богатых почвах лесостепных дубрав под сомкнутым пологом сосновых и еловых культур, что свидетельствует лишь о качестве подстилки, но не почвы, и т. д.).
При выделении типа фитоценозов и насаждений по признаку обилия (господства) отдельных эдификаторов (господствующих видов), коренные и производные ценозы ставят рядом друг с другом, резко отграничивая их как однородные единицы. Между тем насаждения, относящиеся к одному и тому же местообитанию, могут резко изменяться, например, в зависимости от возрастных стадий. Подобные случаи могут вызвать ошибочное отнесение их к разным категориям. Поэтому для более разносторонней и правильной оценки экологии леса целесообразно пользоваться отдельными показателями — видами растений, их ростом, обилием, жизненностью и т. п. признаками, используя ценозы (насаждения) в качестве условных индикаторов, подлежащих экологической расшифровке и уточнению. Геоботанические и таксационные описания насаждений (ценозов) — не более чем сырой материал для сравнительно-экологического анализа и последующей их классификации (определения). Сравнительно-экологическая оценка по индикаторам удобна тем, что она, игнорируя рамки ценозов, дает более точное освещение условий среды и заставляет обращать внимание на ее изменения.
Пользование индикаторами требует разностороннего знания их экологии и биологии. Каждый растительный вид и экотип представляет собой сложное единство, реагирующее на все стороны местообитания. Индикаторная ценность одного и того же вида растений может меняться как объективно, т. е. в зависимости от его поведения, так и субъективно, т. е. в зависимости от того, чем мы в данный момент интересуемся. Самые ценные индикаторы, свидетельствующие о климате, плодородии и увлажнении почв наиболее обобщенно-синтетически, это — древесные породы. Сосна говорит о бедных, ясень — о богатых почвах; кедр и лиственница о холодном климате, черная ольха — об очень влажных почвах и т. д. Однако эти индикаторы не очень чувствительны по отношению к деталям среды. Наиболее чувствительные индикаторы — представители живого напочвенного покрова. В одних случаях они дают нам весьма ценные указания, стоящие на более высоком уровне, чем даже индикаторная ценность древесных пород. В иных случаях, когда виды покрова реагируют на несущественные изменения местообитания коренными изменениями своего состава, ценность их снижается, и они могут служить лишь для оценки менее существенных изменений, зачастую вполне наглядных и очевидных. Например, появление при изреживании насаждения светолюбивых видов свидетельствует об освещенности почвы, но освещенность может быть установлена и без травянистых индикаторов. Однако любая смена коренных ценозов производными все же оставляет нам возможность (а иногда, как например при смене бука и граба более светолюбивыми породами, даже увеличивает возможность) оценивать существенные черты местообитания, отвлекаясь от несущественных сторон в составе и экологии наличных индикаторов.
Подчеркнем, что индикаторами мы называем не только растительные виды (древесные породы всех ярусов насаждения, травянистые, моховые и лишайниковые представители живого напочвенного покрова), но и характерные в экологическом отношении их свойства: присутствие или отсутствие в составе ценоза и его ярусов, обилие, встречаемость, производительность, жизненность. степень развития, возобновляемость, ход сукцессий и т. п. Так, например, отсутствие сосны в естественных коренных насаждениях является одним из ценных индикаторов дубрав.
Появление белоуса на вырубках является характерным признаком влажных и сырых типов леса борового и суборевого ряда, появление овсяницы Беккера — свежего ряда тех же типов и т. п. Произрастание главных древесных пород или их отсутствие, бонитет, возобновляемость, различные особенности их биологии и экологии — наиболее точные измерители среды.
Техническая сторона методики сравнительной экологии заключается в составлении экологических рядов, представляющих собой ряды древостоев — ценозов, расположенных по признакам нарастания или убывания количества какого-либо одного ведущего экологического фактора. Основанием для построения экологических рядов является конкретный типологический профиль того или иного лесного массива, отображающий его основное разнообразие. Вдоль профиля производят съемку рельефа и подробное таксационное и геоботаническое описание насаждений. Экологические ряды строятся таким образом, что весь типологический профиль или отдельные его части представляются на диаграмме в виде более или менее чистого ряда типов леса, выделенных по признакам растительности. Внешние признаки местообитания могут служить в этом случае лишь корректирующей придержкой. В экологическом ряду могут быть поставлены по соседству типы леса, территориально разобщенные, имеющие местообитания, не сходные по внешним признакам, но обязательно близкие по экологии.
Третьим техническим приемом сравнительной экологии является составление индикаторных спектров, которые представляют собой перечень растений с характеристикой их обилия, а при необходимости — мощности развития, жизненности и других интересных в данном случае свойств. Для древесных пород указывают их долю в составе насаждения, бонитет и другие лесоводственные и биологические признаки. Индикаторные спектры составляются для каждого экологического ряда по каждому ведущему фактору в отдельности: трофности, увлажнению, тепловым условиям, освещению, степени влияния скотосбоя и т. п.
В качестве основы для типологического профиля иногда используют внешние признаки местообитания, при этом отводят насаждениям их корректирующую, сопутствующую роль. Сюда относится, например, геоморфологический профиль через речную долину лесостепной зоны, служивший Г. Ф. Морозову основой для рассмотрения разнообразия лесов этой зоны.
Основой экологического ряда является растительность как отражение местообитания. Но при составлении экологических рядов иногда обходятся без углубления в отдельные стороны местообитания и его ведущих факторов. Составление индикаторных спектров способствует познанию причин, обусловливающих характер тех или иных местообитаний, или их отдельных свойств.
При изучении экологического разнообразия лесов в каждом лесном массиве подыскивают короткий типологический профиль (ряд, комплекс), составленный из всех или большинства встречающихся здесь типов леса. Более или менее полно совпадающие с экологическими рядами конкретные типологические профили являются как бы живыми диаграммами. Они дают возможность сравнивать типы леса на основании, гарантирующем от случайностей, и более обстоятельно изучать переходы между ними. Переходы очень важны, так как с их помощью удается установить степень экологического родства и различия между изучаемыми лесами.
Сказанное выше предполагает, что в один и тот же тип местообитания могут входить почвы, разные по внешним (морфологическим) признакам. Так и бывает в природе. Например, сухие и свежие боры встречаются на глубоких песках и на мелких скелетных, суглинистых по составу мелкозема почвах. Субори встречаются не только на глубоких, однородных супесях, но и на песках, подстилаемых на глубине 2—3 м суглинками, или, наконец на песках, переслоенных супесчаными или суглинистыми прослойками. Дубравы и рамени распространены не только на плодородных суглинистых почвах, но и на песках или супесях, неглубоко подстилаемых суглинками, и т. д. Обычные почвенные описания и карты имеют большую ценность для полеводства, но в лесном хозяйстве и лесоустойстве незаменимыми являются типы лесов и лесных местообитаний, определяемые на основе растительности. Региональные почвенные исследования имеют ценность лишь как дополнение к типологическим исследованиям при условии тесной привязки и полном подчинении типов почв типам леса — местообитания.
Относящиеся сюда вопросы более конкретно и подробно были обсуждены в интересной дискуссии, разгоревшейся на страницах журнала «Лесное хозяйство» (1939, № 11 и 1940 № 6) между проф. В. Н. Сукачевым, с одной стороны, проф. В. Э. Шмидтом и доц. П. Г. Кроткевичем — с другой. Первый ученый стоял на точке зрения, утверждавшей, что индикаторные свойства растений несовершенны и что лучшим показателем плодородия почв является исследование самой почвы. В. Н. Сукачев допускал суждение о плодородии почв по растительности лишь как временную меру, применяемую при отсутствии более совершенных методов почвенного анализа. Такую же точку зрения развивал и последователь В. Н. Сукачева А. П. Петров («Советская ботаника», 1947, № 1); эту теорию критиковал П. С. Погребняк (Основы лесной типологии, Киев, 1955, стр. 78—79). В. Э. Шмидт и П. Г. Кроткевич представляли себе внутренний смысл дискуссии в решении вопроса, чем же является почвенное плодородие? На него они отвечали с полным основанием: плодородие почвы есть отношение растения к почве; растение (точнее — урожай растения) является единственным абсолютным критерием плодородия. Точку зрения, представляющую себе более совершенным суждение о плодородии по признакам почвы, они считали ошибочной. Таких же взглядов придерживался и Л. Г. Раменский, для которого растительность была «высшей инстанцией» оценки почвенного плодородия.