Выше была рассмотрена роль бактериофага лишь как переносчика наследственного материала от одной клетки к другой при трансдукции.
Но фаги интересны не только как пассивные переносчики наследственного материала. Сами по себе они так же, как й вирусы высших организмов, является генетическим объектом. Гены вирусов могут претерпевать мутации и рекомбинации, спонтайные и. индуцированные различными воздействиями: ультрафиолетовыми лучами, ионизирующими излучениями и химическими агентами. Мутации могут касаться таких признаков фага, как скорость лизирования бактериальной клетки, способность заражать бактерии определенного штамма, антигенная природа белковой оболочки фага и т. д. Эти мутантные свойства фагов могут сохраняться длительно е время при их размножении.
При заражении клетки вирулентным фагом биосинтез клетки, находившейся под контролем генома хозяина, останавливается: синтез ДНК бактерии полностью прекращается, происходит обновление РНК и соответственно синтезируются новые белки для построения фаговых частиц. Вновь синтезируемые молекулы ДНК, РНК и белка на этом этапе осуществляются под контролем ДНК фага. При этом если геном бактерии не способен синтезировать какую-либо аминокислоту, а геном фага обладает такой возможностью, то геном фага может обеспечить ее синтез. Геном фага настраивает биосинтез на свой лад. В данном случае мы вправе говорить о генетическом паразитизме.
Фаговая ДНК сначала в клетке размножается и находится в виде отдельных нитей (вегетативный фаг). В это же время гены фага контролируют формирование соответствующей белковой оболочки и части зрелой частицы. ДНК зрелых фаговых частиц в клетке хозяина не воспроизводит себя.
Примером генетического анализа фага может служить следующий опыт. Существуют фаги (r) с мутантным фактором, обусловливающим появление на культуре Escherichia coli больших светлых пятен (зон лизиса или подавления роста бактерий), и фаги с мутантным фактором (m), которые обусловливают появление мелких зон лизиса. Культуру кишечной палочки заражали одновременно двумя мутантными штаммами фагов r—m+ и г+m—. При такой смешанной инфекции, когда с большой вероятностью все клетки были инфицированы указанными двумя генотипами фагов, появлялись четыре типа пятен: большие светлые, маленькие темные, маленькие светлые и большие темные. Первые два соответствовали исходным мутантным частицам фагов r—m+ и г+m—, а два последних, очевидно, имели новое сочетание генов: r—m— и г+m+. Они могли возникнуть лишь в результате рекомбинации генов двух исходных штаммов фага, произошедшей в бактериальной клетке.
Картирование генов по трем точкам, примененное Т. Морганом для дрозофилы, было использовано и для составления генетической карты у фага Т4. Так, в таблице показано сцепление генов в гибридном скрещивании и рекомбинация у фага Т4, а на рисунке — генетическая карта этого фага.

Сцепление генов при тригибридном скрещивании и рекомбинация у фага T4
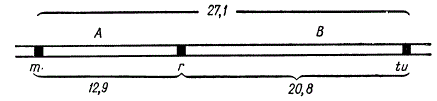
Картирование по трем точкам у фага Т4 — генам m, r, tu
Гены фага Т4 располагаются на генетической карте линейно. При изучении тесно сцепленных генов здесь обнаруживается отрицательная интерференция; один гецетический обмен повышает вероятность другого обмена на соседнем участке. В силу этого сумма частот рекомбинаций в двух соседних участках будет всегда выше, чем между двумя крайними генами. Однако здесь учитывается рекомбинация неоднократной репликации, поскольку в процессе размножения фага в клетке хозяина происходит многократное воспроизведение генома. При размножении фага в клетке возникают фаги разной генетической структуры, между которыми могут происходить неоднократные «скрещивания» и репликации. Следовательно, процесс рекомбинации в опытах с фагами отражает процесс рекомбинации в размножающейся популяции фагов внутри хозяина-клетки, и внутри одной зараженной бактерии нельзя получить равенства реципрокных комбинаций фагов. В результате одного события образуются нереципрокные рекомбинации, что и отличает фаговую рекомбинацию от истинной хромосомной.
Равенство реципрокных комбинаций может быть достигнуто только при исследовании большого числа зараженных разных бактерий одним и тем же штаммом фага.
Каким же образом может происходить рекомбинация фагов внутри клетки бактерии? Поскольку «хромосома» фага имеет «чистую» молекулу ДНК, то скрещивание должно осуществляться на молекулярном уровне между молекулами ДНК в момент репликации.
Одной из гипотез, объясняющих рекомбинацию у фагов, является гипотеза copy-choice, или partial-replica. Смысл этой гипотезы заключается в том, что при репликации по оси матрицы ДНК происходит смена матриц с одной нити ДНК на другую. Сначала репликация идет по одной матрице, но вследствие повреждения участка на пути репликации данной матрицы дальнейшая репликация молекулы ДНК может идти по другой ДНК — неповрежденной. Подтверждением этой гипотезы служит тот факт, что под влиянием мутагенов частота рекомбинаций у фагов повышается.
В объяснении процесса рекомбинации фагов с точки зрения указанной гипотезы много трудностей, хотя самое явление рекомбинации наследственных фагов не вызывает сомнения.
Генетика вирусов приобретает сейчас исключительно важное значение для изучения роли нуклеиновых кислот в синтезе белка, а также для познания химических основ наследственности.
Генетические исследования на микроорганизмах и вирусах раскрыли новый мир явлений и огромные перспективы в деле управления наследственной природой организмов. Такие явления, как трансформация, трансдукция, половой процесс у бактерий, мутации рекомбинации у вирусов, значительно расширили наши представления о материальных носителях наследственности и многообразии механизмов их передачи. Благодаря этим явлениям установлено, что:
1) ДНК участвует в передаче наследственной информации при клеточном делении;
2) процессы мутаций и рекомбинаций, установленные ранее для высших организмов, являются общими и для микроорганизмов;
3) у эукариотов процесс рекомбинации, как правило, является Реципрокным; он осуществляется при половом размножении в мейозе путем внутрихромосомной рекомбинации, а при вегетативном размножении — путем митотической рекомбинации в митотическом цикле;
4) у прокариотов (например, фаговых частиц) процесс рекомбинации в силу особого их размножения, как правило, нереципрокный; у микроорганизмов часто наблюдается односторонний и частичный обмен генетическим материалом.