Изучение явления трансформации послужило толчком к открытию другого явления — трансдукции — переноса и рекомбинации генов у бактерий с помощью бактериофага.
Опыт, позволивший открыть этот новый генетический механизм и новый способ изучения наследственности, заключается в следующем.
U-образная трубка в нижней части была разделена посредине бактериальным фильтром. В одну половину этой трубки были помещены тифозные бактерии (Salmonella typhimurium) штамма 22А, а в другую половину трубки — штамма 2А. При этом бактериальные клетки не могли переходить сквозь перегородку.
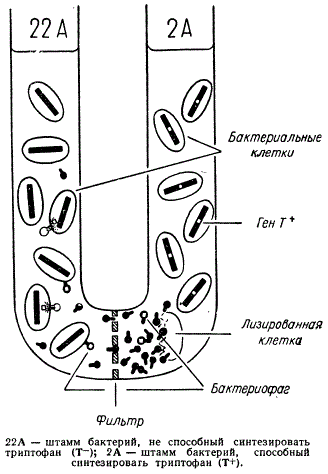
Схема опыта, демонстрирующего явление трансдукции у Salmonella typhimurium
Штамм 22А нес мутацию, блокирующую синтез триптофана Т—, и поэтому при культивировании бактерии нуждались в добавке триптофана в среду. Штамм бактерии 2А имел мутацию, блокирующую синтез гистидина Н—, и поэтому нуждался в нем при культивировании.
После инкубации этих двух разных штаммов в трубке, разделенной только бактериальным фильтром, был произведен рассев клеток обоих штаммов. При рассеве клеток штамма 22А на среде, лишенной триптофана, было обнаружено небольшое число колоний. Следовательно, некоторые клетки штамма 22А каким-то образом приобрели способность синтезировать триптофан и смогли дать колонии на среде без этой аминокислоты. Частота появления таких клеток была равна 1х10-5.
Можно было предположить, что эти измененные клетки что явились или в результате обратной мутации от Т— к Т+ или перехода трансформирующего фактора от штамма 2А. Но штамм 22А отличался высокой стабильностью, и поэтому указанную частоту появления (106) клеток генотипа Т+ нельзя было объяснить возникновением обратных мутаций. Трансформирующий фактор в среде также не был обнаружен. Фильтрующимся агентом, переносящим ген Т+ от штамма 2А к штамму 22А, оказался бактериофаг.
Таковы первые факты, доказавшие передачу наследственной информации с помощью бактериофага от бактерии одного генотипа к бактерии другого генотипа. Это открытие было сделано в 1952 г. Н. Циндером и Дж. Ледербергом.
Используемый в исследованиях Циндера и Ледерберга штамм 22A Solmonella typhirnurium не обладал способностью синтезировать триптофан, но после совместного содержания в разделенной фильтром U-образной трубке со штаммом 2А приобрел свойство синтезировать триптофан. Это могло произойти только в том случае, если фаг, вышедший из клеток штамма 2А, проник через фильтр, внедрился в некоторые клетки штамма 22А и передал им часть наследственной информации — фрагмент наследственного материала штамма 2А.
Следовательно, ДНК фага, лизогенизирующего бактерию, каким — то образом претерпевает рекомбинацию с ДНК бактериальной клетки, в силу чего в новые фаговые частицы включаются гены клетки-хозяина. Эти фаги, заражая вновь клетки другого генотипа, также передают ей свою ДНК с новой информацией. Так, клетки штамма 22А приобрели ген, ответственный за синтез триптофана.
Как мы видели, фаги являются переносчиками наследственной информации от бактерии одного генотипа к бактерии другого генотипа. И это возможно только при условии, если ДНК фага вступает интимные связи с ДНК хромосом бактериальных клеток. Это явление переноса отдельных наследственных задатков от бактерии-донора, в которой происходили размножение фага и рекомбинация Готического материала фага и хозяина-бактерии к реципиенту, и называется трансдукцией.
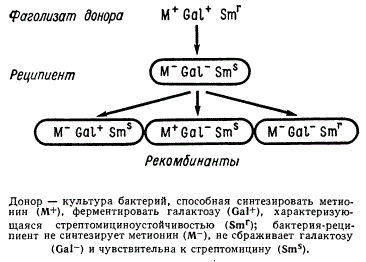
Схема генетической трансдукции
Донором является культура бактерии, способная синтезировать метионин М+ и ферментировать галактозу Gal+, а также имеющая стрептомициноустойчивость Smr. Бактерия-реципиент не синтезирует метионин М—, не сбраживает галактозу Gal— и чувствительна к стрептомицину Sms. Фаголизат, полученный от донора M+Gal+Smr, вносят в культуру реципиента M—Gal—Sms. После инкубации клетки реципиента рассевают на соответствующих селективных средах, в результате чего обнаруживаются три новых класса рекомбинантов M—Gal+Sms, M+Gal—SmS, M—Gal—Smr.
В случае трансдукции донор через фаг передает лишь отдельный фрагмент ДНК. Поэтому инфицированные бактерии реципиента являются как бы диплоидными по переданному фрагменту (мерозиготы) и частично гетерозиготами (гетерогеноты), в потомстве которых могут быть рекомбинантные бактерии M+Gal—Sms и M—Gal—Sms возникшие при трансдукции.
Судьба переданного фрагмента хромосомы донора в клетке реципиента может быть различной. Этот фрагмент может, во-первых, внедриться в хромосому хозяина и реплицироваться совместно и синхронно с соответствующим участком хромосомы хозяина (завершенная трансдукция), во-вторых, может быть удален из клетки хозяина и, в-третьих, может сохранять автономность и передаваться от клетки к клетке независимо от хромосомы хозяина (абортивная трансдукция).
Фаг может переносить самые различные гены бактерий, обусловливающие определенный характер синтеза аминокислот, различные ферментативные свойства, устойчивость к антибиотикам (стрептомицину, пенициллину) и иммунность к другому фагу. Как правило, одновременно трансдуцируется один, реже — два тесно сцепленных гена и очень редко три гена. Эта особенность была использована в опытах М. Демереца с сотрудниками, которым удалось посредством учета результатов трансдукции провести картирование тесно сцепленных генных локусов, обеспечивающих синтез цистеина у Salmonella.
Таким образом, трансдукция так же, как и трансформация, является своеобразным процессом рекомбинации генов. Рекомбинация генов является одним из механизмов, осуществляющих у бактерий комбинативную изменчивость, которая у высших организмов обеспечивается мейозом.