Учет количества возникающих мутаций необходим при исследовании природы гена, его изменения, для понимания механизма влияния внешних условий и физиологического состояния организма на мутационный процесс.
Однако трудность подобного учета заключается в том, что часто не удается отличить мутацию от рекомбинации. Как мы видели в результате процессов расщепления и кроссинговера, в потомстве могут появляться новые наследственные признаки, которые легко принять за мутации. Такое же явление возможно при взаимодействии генов; рекомбинации могут имитировать мутации особенно в тех случаях, когда гены тесно сцеплены и редко разделяются в результате кроссинговера.
Методы обнаружения мутаций должны быть разными в зависимости от особенностей объекта — главным образом способа размножения организма. При вегетативном и бесполом размножении многоклеточных организмов мутации учитываются в соматических тканях, дающих побег, в потомстве одной особи (по клонам). У самооплодотворяющихся растений и животных рецессивные мутации проявляются в следующем же после возникновения мутации поколении перекрестно оплодотворяющихся организмов возникающие мутации переходят в гетерозиготное состояние в популяции особей, для выявления их необходимо применять близкородственное скрещивание (инбридинг), чтобы увеличить вероятность встречи особей, несущих в себе мутировавший ген.
Как уже говорилось выше, объективно регистрировать общее число возникающих мутаций пока практически невозможно. Однако подсчет мутаций в отдельных локусах и определенного типа мутаций вполне доступен. Некоторые видимые морфологические изменения можно учитывать довольно точно; несколько более сложным является определение физиологических и биохимических изменений у многоклеточных организмов. Последнее удается лишь при применении стандартных тестов на определенный химический состав или физиологическую реакцию организма и т. д., т. е. по ответу «да» или «нет».
Легче всего обнаруживаются видимые доминантные мутации, которые могут проявляться в гетерозиготном состоянии в первом же поколении, труднее анализировать рецессивные мутации. Для выявления последних требуется специальный генетический анализ в ряду поколений. Для того чтобы учитывать мутации, особенно рецессивные, возникшие как единичные изменения в хромосомах половых клеток, их необходимо переводить в гомозиготное состояние. Для дальнейшего анализа мутантную линию скрещивают с линией-анализатором, имеющей одну или несколько маркированных групп сцепления. Такой подход позволяет не только подтвердить ее наследование, но и сберечь время на анализ принадлежности мутации к соответствующей группе сцепления.
Для хорошо изученных в генетическом отношении объектов (дрозофила, кукуруза, ряд микроорганизмов) с установленными группами сцепления изучение новой мутации проводить довольно легко, действуя только что указанным путем. Для этих же объектов разработаны специальные методики учета частоты мутаций, возникающих в отдельных хромосомах. Так, например, для обнаружения видимых мутаций в половой хромосоме у дрозофилы используется методика сцепленных Х-хромосом — уу (двойной желтый). По данной схеме скрещивания можно обнаружить отдельные видимые рецессивные сцепленные с полом мутации, возникшие в Х-хромосоме половых клеток отцовского организма и проявляющиеся у мужского пола в F1.
Наиболее объективно можно учитывать рецессивные летальные мутации, приводящие в гомозиготном состоянии к смерти несущих их особей. Для учета таких мутаций в половой хромосоме дрозофил, Г. Мёллером была разработана методика СIВ.
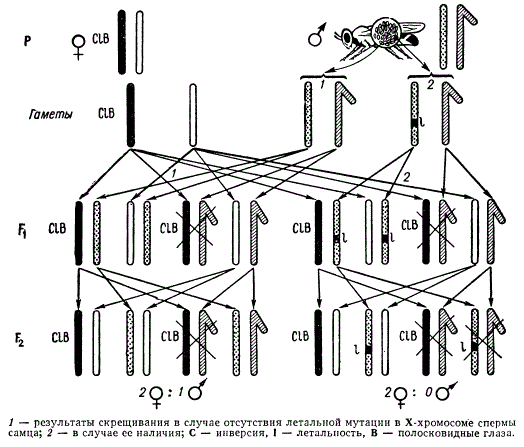
Метод обнаружения рецессивных сцепленных с полом летальных мутаций у дрозофилы
Генетическая структура линии СIВ характеризуется тем, что одна из Х-хромосом самки маркирована доминантным геном Bar (полосковидные глаза). В этой же хромосоме имеется инверсия, обозначаемая буквой С. Эта инверсия препятствует кроссинговеру и обладает рецессивным летальным эффектом 1, т. е. зиготы, несущие две такие Х-хромосомы, погибают. Этими тремя начальными буквами (СIВ) и обозначена линия-анализатор на летальные мутации в половых хромосомах дрозофилы.
В случае если в одном из спермиев анализируемого самца в Х-хромосоме возникает летальная мутация, то при оплодотворении таким спермием яйцеклетки с Х-СIВ-хромосомой развивается гетерозиготная по данной летали самка. В F1 будут встречаться самки двух типов: с нормальными (круглыми) и с полосковидными глазами (Bar). Каждая из этих самок несет по одной Х-хромосоме отца. Самцы, несущие хромосому С1В, не развиваются, так как у них деталь находится в гемизиготном состоянии. Поэтому самцы в F1 встречаются только с нормальными глазами. Для дальнейшего анализа самок F1 с геном Bar скрещивают индивидуально с нормальными самцами (каждая пара в отдельной пробирке).
Если самка F1 получила Х-хромосому анализируемого самца с летальной мутацией и, следовательно, стала гетерозиготной по ней, то в потомстве F2 такой самки не появится самцов, так как все они погибнут. В результате расщепление по полу в такой культуре оказывается в отношении 2♀:0♂ (причем половина самок с нормальными глазами, а половина — с полосковидными). Очевидно, что число исследованных на обнаружение летальных мутаций самок СIВ из F1-дочерей одного исходного анализируемого самца будет соответствовать числу исследованных Х-хромосом в его сперматозоидах.
Хотя эта методика и очень удобна для количественного учета возникающих летальных мутаций, но у самок F1 между половыми хромосомами иногда может происходить двойной перекрест, что приводит к снижению истинной частоты летальных мутаций.
В настоящее время для анализа частоты возникновения летальных мутаций в Х-хромосоме самцов применяется другая методика, названная Меллер-5, или М-5.
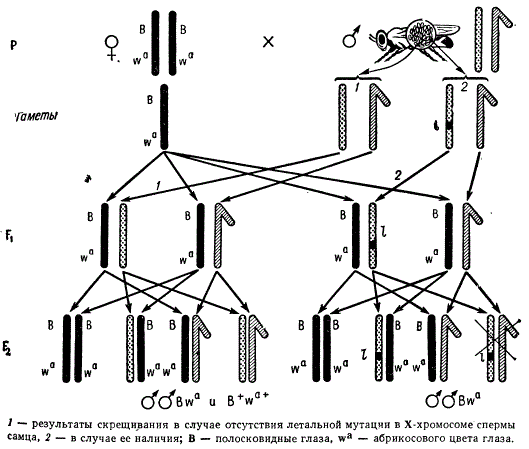
Метод обнаружения рецессивных сцепленных с полом летальных мутаций у дрозофилы (метод М-5)
В этой методике применяется линия-анализатор. Преимущество такой линии-анализатора заключается в том, что обе Х-хромосомы самки содержат по две Инверсии, не связанные с летальным действием. В силу наличия двух инверсий перекрест между хромосомами более затруднен. Кроме того, обе хромосомы самки маркированы тремя генами: sc8, В, wa. Самцы в этой линии жизнеспособны. При анализе самца дикого типа методом М-5 мы получаем в F2 по два фенотипических класса самок и самцов. Если же в анализируемой Х-хромосоме исследуемого самца возникла летальная мутация, то в F2 будут самцы одного фенотипического класса: sc8, В, wa, а самцы дикого типа не появятся. При этом каждая индивидуальная культура F2, являющаяся потомством лишь одной самки F1 соответствует одной исследованной Х-хромосоме самца Р.
Для учета летальных мутаций в аутосомах дрозофилы существует другая методика. В этом случае учет возникающих рецессивных летальных мутаций производится в F3.
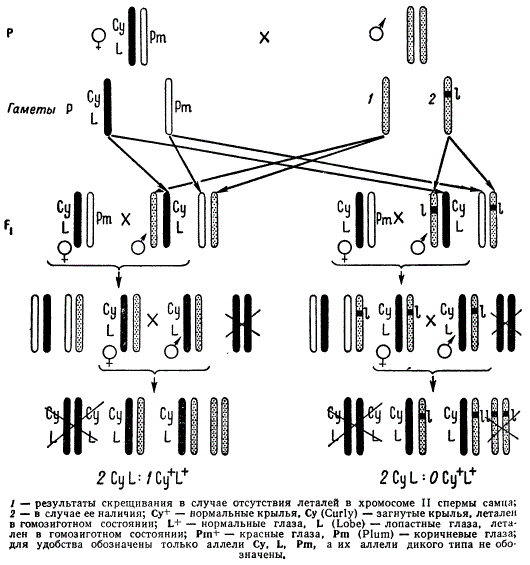
Метод обнаружения летальных мутаций в аутосомах дрозофилы (метод сбалансированных леталей CyL/Pm)
Для проявления летальной рецессивной мутации в аутосоме необходимо, чтобы мутация оказалась в обеих испытываемых хромосомах, т. е. в гомозиготном состоянии. Обычно для этой цели у дрозофилы используют специальную линию, позволяющую учитывать летальные мутации во второй паре аутосом. В этой линии одна из хромосом содержит две большие инверсии — по одной в каждом плече, доминантные гены Cy (Curly — загнутые крылья) и L (Lobe — маленькие глаза), каждый из которых в гомозиготном состоянии вызывает летальный эффект; гомологичная хромосома, также содержащая инверсию, маркирована геном Pm (Plum — коричневатые глаза). Этот метод обнаружения мутаций иногда называют методом CyL/Pm.
Испытываемого самца скрещивают с самкой из линии CyL/Pm. В F1 все мухи CyL являются гетерозиготными по той или другой гомологичной хромосоме II анализируемого самца. Из F1 выбирают самцов CyL, поскольку у них кроссинговер подавлен. Каждого такого самца, несущего по одной анализируемой хромосоме, индивидуально скрещивают с самкой исходной линии CyL/Pm. В F2 половина мух (самцов и самок) в каждой культуре оказывается гетерозиготной по одной и той же анализируемой хромосоме. Фенотипически у этих мух проявляются только гены Су и L. Таких самок и самцов скрещивают между собой. В случае отсутствия летальной мутации в анализируемой хромосоме происходит расщепление в отношении 2CyL : 1Cy+L+. Данное отношение возникает в силу того, что гомозиготные мухи CyL не выживают. Если в анализируемой хромосоме имеется деталь, то в культуре F3 будут мухи одного фенотипа — CyL, а нормальные по фенотипу будут погибать в силу гомозиготного состояния рецессивной летальной мутации.
Таким методом можно учитывать частоту рецессивных летальных мутаций во второй хромосоме дрозофилы.
Вообще летальные мутации по генетической природе являются смешанным типом мутаций, к которым относятся как различного рода хромосомные перестройки, так и изменения отдельных генов.
В итоге рассмотрения общей характеристики мутационного процесса мы приходим к следующим выводам.
- Генные мутации являются скачкообразными изменениями отдельных локусов хромосом — генов.
- Мутантные гены сохраняют свойство репродукции при делении ядра клетки, вследствие чего мутационные изменения наследуются.
- Мутации могут быть прямыми и обратными.
- Частота мутирования в обоих направлениях характерна для каждого локуса.
- Спонтанный мутационный процесс обусловливается свойством самого гена, системой генотипа, физиологическим состоянием организма и колебанием факторов внешней среды.
- Каждый локус — ген может мутировать в несколько состоянии, образуя серию множественных аллелей.
- Мутации у различных видов организмов образуют гомологические ряды наследственных изменений.