Флоэма и ксилема обычно сопровождают друг друга, образуя проводящие пучки.
По типу элементов, входящих в состав флоэмы и ксилемы, проводящие пучки получили еще название сосудисто-волокнистых. В молодых органах большинства растений сосудисто-волокнистые пучки идут раздельно. На более поздних стадиях у двудольных и голосеменных растений проводящие пучки сливаются, образуя оплошной цилиндр, состоящий из тканей древесины и луба. У однодольных пучковая структура сохраняется на всех стадиях развития органов. Сосудисто-волокнистые пучки возникают из меристемы, называемой прокамбием. Прокамбий состоит из удлиненных, очень тонкостенных клеток с заостренными концами. Он закладывается недалеко от точки роста органа, в свою очередь, дифференцируясь из промеристемы. В стебле прокамбий возникает в виде небольших участков, расположенных кольцом. Когда таких участков образуется мало, то промежутки между ними больше, а при значительном числе участков прокамбия или при увеличении их размеров по периферии органа расстояния между отдельными участками могут стать очень незначительными. На рисунке изображены участки прокамбия в поперечном, а на другом рисунке — в продольном разрезах. Клетки прокамбия отличаются от окружающих клеток более густым и зернистым содержимым, они узкие и вытянуты в продольном направлении.
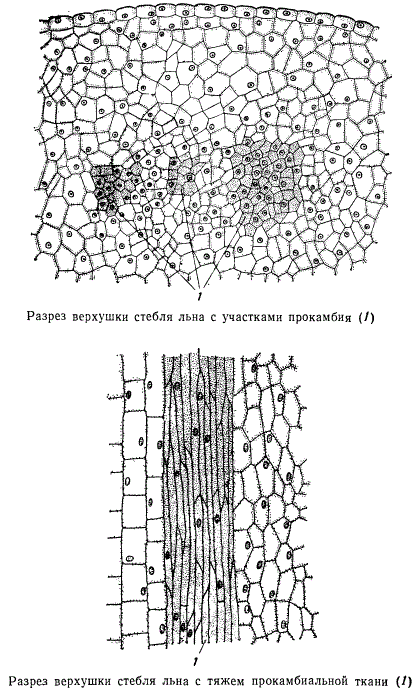
Первыми в прокамбии дифференцируются элементы флоэмы, называемые протофлоэмой. Первые по времени возникновения сосуды называют протоксилемой. Образовавшиеся из прокамбия сосудисто-волокнистые пучки состоят из флоэмы и ксилемы и называются первичными. Кроме протофлоэмы и протоксилемы, среди первичных элементов проводящей ткани различают метафлоэму и метаксилему. Возникновение этих элементов представляет собой вторую ступень дифференциации прокамбия. Первичная флоэма сформирована ситовидными трубками со спутниками, лубяными волокнами и лубяной паренхимой. У некоторых растений, например, у виноградной лозы, ситовидные трубки первичной флоэмы имеют горизонтально расположенные ситовидные пластинки с одним большим ситом, тогда как у более поздно возникающих ситовидных трубок вторичной флоэмы ситовидные пластинки косые, с несколькими ситами. Первичная ксилема состоит из сосудов и трахеид, для которых характерны кольчатые и спиральные утолщения оболочек.
В пучках наиболее распространенного типа на поперечных срезах флоэма и ксилема располагаются по одному радиусу и дифференцируются из клеток прокамбия навстречу друг другу. Между дифференцированными элементами остается слой меристематических клеток.
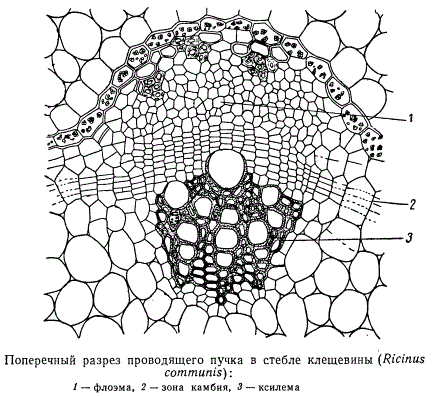
Деления в этом слое вскоре становятся упорядоченными: клетки начинают делиться главным образом тангентальными перегородками. Формирующаяся таким образом меристематическая ткань отличается по морфологии клеток и характеру делений от прокамбия и носит название камбия. Клетки камбия дают начало элементам флоэмы и ксилемы, называемым вторичной флоэмой и вторичной ксилемой. Элементы вторичных проводящих тканей имеют более сложную организацию, чем соответствующие элементы первичных тканей сосудисто-волокнистых пучков.
Камбий как меристематическая ткань несколько отличается от прокамбия по структуре оболочек (оболочки более плотные, с порами), а также большей вакуолизацией цитоплазмы. Клетки камбия прозенхимные, сильно вытянуты вдоль оси органа. Например, длина камбиальных клеток в стволе яблони в среднем в 25 раз больше размеров их поперечников. Особенно длинны камбиальные клетки у хвойных и вообще голосеменных. У сосны длина камбиальных клеток превышает ширину больше чем в сто раз. Наоборот, у травянистых растений камбиальные клетки относительно короткие.
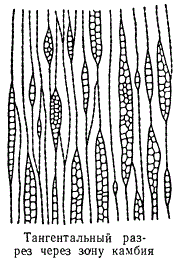 Специальными измерениями установлено, что длина камбиальных клеток у древесных двудольных колеблется в пределах от 150 до 600 мк (может быть, и немного больше), а у хвойных достигает 5000 мк (лиственница). Если через камбиальную зону сделать тангентальный разрез, то можно убедиться, что камбиальные клетки на концах заострены. На радиальных же разрезах камбиальные клетки имеют вид удлиненных прямоугольников. На тангентальных разрезах через камбиальную зону видно, что не все меристематические клетки в этой зоне одинаковы. Одни клетки вытянуты, веретенообразны, другие округлы, как паренхимные клетки. Первые — настоящие камбиальные клетки — дают начало прозенхимным анатомическим элементам: ситовидным трубкам, членикам сосудов, трахеидам, веретеновидным компонентам древесинной паренхимы и пр. Мелкие же паренхимоподобные клетки камбиальной зоны образуют клетки сердцевинных лучей.
Специальными измерениями установлено, что длина камбиальных клеток у древесных двудольных колеблется в пределах от 150 до 600 мк (может быть, и немного больше), а у хвойных достигает 5000 мк (лиственница). Если через камбиальную зону сделать тангентальный разрез, то можно убедиться, что камбиальные клетки на концах заострены. На радиальных же разрезах камбиальные клетки имеют вид удлиненных прямоугольников. На тангентальных разрезах через камбиальную зону видно, что не все меристематические клетки в этой зоне одинаковы. Одни клетки вытянуты, веретенообразны, другие округлы, как паренхимные клетки. Первые — настоящие камбиальные клетки — дают начало прозенхимным анатомическим элементам: ситовидным трубкам, членикам сосудов, трахеидам, веретеновидным компонентам древесинной паренхимы и пр. Мелкие же паренхимоподобные клетки камбиальной зоны образуют клетки сердцевинных лучей.
Полагают, что собственно камбий состоит только из одного слоя клеток. Все прочие слои камбиальной зоны — молодые, еще окончательно не оформившиеся элементы флоэмы и ксилемы. Из того факта, что древесина во всех органах значительно массивнее лубяной части, можно судить о том, что работа камбия по откладыванию элементов ксилемы и флоэмы неравномерна.
Камбиальные клетки раза в четыре чаще превращаются в элементы ксилемы, чем в элементы флоэмы.
Если подсчитать число веретенообразных камбиальных клеток и образовательных клеток сердцевинных лучей на поперечных разрезах стеблей различного возраста у одного и того же растения, можно легко убедиться, что числа эти с возрастом растения весьма заметно увеличиваются. Так, однолетний стебель веймутовой сосны имеет веретенообразных камбиальных клеток — 724, камбиальных клеток сердцевинных лучей — 70, стебель же в возрасте 60 лет — соответственно 23100 и 8796.
Часть камбиальной ткани, образуемая клетками сердцевинных лучей, по объему значительно меньше той части, которая состоит из веретенообразных камбиальных клеток.
Например, у 60-летней сосны на долю камбиальных клеток сердцевинных лучей приходится приблизительно 1/8 всей периферии камбиального кольца. У многих двудольных длина окружности, занимаемой камбиальными клетками сердцевинных лучей, нередко относительно выше, чем у хвойных, но все же чаще всего суммарная длина дуг, состоящих из веретенообразных камбиальных элементов, превосходит общую длину дуг, слагаемую из камбиальных клеток сердцевинных лучей. Камбиальные клетки делятся регулярно. Преобладают периклинальные деления, в результате которых образуются инициальные клетки элементов флоэмы (кнаружи от камбия) и элементов ксилемы (внутрь от камбиального слоя).
В осевых органах (стебле, корне) камбий образует обычно сплошное кольцо. Увеличение слоев клеток, откладываемых камбием, а также разрастание в поперечнике многих образованных им элементов обусловливает равномерный рост органов в толщину. В соответствии с увеличением радиуса осевого органа длина окружности, по которой располагаются клетки камбиального слоя, также увеличивается. Это увеличение происходит за счет радиальных делений клеток камбия.
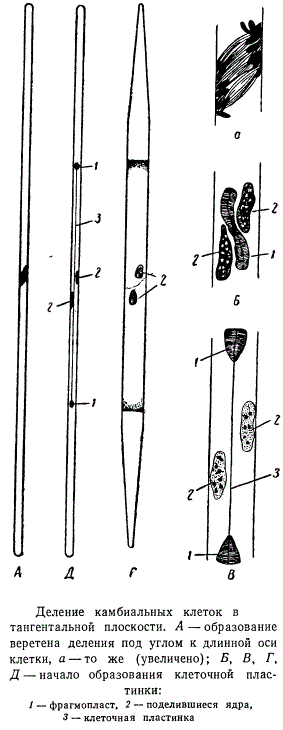 Деление радиальными перегородками приводит к увеличению числа камбиальных клеток и разрастанию камбиального пояса по периферии органа. Деление клеток камбия тангентальными перегородками создает инициальные клетки флоэмы и ксилемы. Остановимся более подробно на тангенталыных делениях. В связи с узким просветом камбиальной клетки при делении ядра плоскость деления располагается косо по отношению к продольной оси клетки. В цитокинезе происходит некоторое изменение положения плоскости деления. На рисунке видно, что фрагмопласт, расположенный между дочерними ядрами, несколько изогнут. После того как образуется центральная часть клеточной пластинки, плоскость деления выравнивается, проходя через продольную ось клетки. Постепенно клеточная пластинка формируется вдоль всей плоскости деления, разделяя протопласт на две равные части, из которых каждая содержит по одному дочернему ядру. Оба ядра остаются в центре своей клетки в продолжение всего процесса образования клеточной пластинки.
Деление радиальными перегородками приводит к увеличению числа камбиальных клеток и разрастанию камбиального пояса по периферии органа. Деление клеток камбия тангентальными перегородками создает инициальные клетки флоэмы и ксилемы. Остановимся более подробно на тангенталыных делениях. В связи с узким просветом камбиальной клетки при делении ядра плоскость деления располагается косо по отношению к продольной оси клетки. В цитокинезе происходит некоторое изменение положения плоскости деления. На рисунке видно, что фрагмопласт, расположенный между дочерними ядрами, несколько изогнут. После того как образуется центральная часть клеточной пластинки, плоскость деления выравнивается, проходя через продольную ось клетки. Постепенно клеточная пластинка формируется вдоль всей плоскости деления, разделяя протопласт на две равные части, из которых каждая содержит по одному дочернему ядру. Оба ядра остаются в центре своей клетки в продолжение всего процесса образования клеточной пластинки.
Хотя начальные камбиальные клетки в зрелом стебле покрытосеменных в среднем значительно короче камбиальных клеток голосеменных, основные черты клеточного деления у них одинаковы. У голосеменных и примитивных двудольных (древесные представители порядка лютикоцветных), имеющих в ксилеме только трахеиды, веретенообразные камбиальные клетки, никогда не распределяются совершенно правильными рядами, тогда как у высокоорганизованных двудольных камбиальные клетки этого типа образуют на тангентальных срезах параллельные ряды. Камбий второго типа называют ярусным. Особенности расположения составляющих его клеток определяются строго упорядоченным характером клеточных делений. В ярусном камбии бывают только тангентальные и радиальные деления, поперечных и косых делений здесь не наблюдается. В камбиальной ткани, характеризующейся отсутствием строгой ярусности в расположении клеток, наряду с типичными тангентальными и радиальными делениями происходят также деления поперечно-косыми перегородками.
Активность камбия в течение всей жизни растения не остается постоянной. Существует сезонная ритмика камбиальной деятельности у древесных и кустарниковых пород. Сезонные изменения активности камбия сопровождаются определенными изменениями в морфологии камбиальных элементов. Так, отмечаются сезонные изменения размеров ядер камбиальных клеток. Например, у веймутовой сосны покоящиеся ядра веретенообразных камбиальных клеток в период опадения хвои и зимой заметно длиннее и уже, чем весной и летом. Особенно четко это различие в морфологии покоящихся ядер выражено в камбиальных клетках корней.
Деятельность камбия по своей активности неодинакова не только в различные времена года у растений одного и того же вида, но даже у одного и того же растения в каждый данный момент в различных его участках. Например, в одних участках стебля камбиальные клетки могут находиться в состоянии активного деления, а в соседних участках того же стебля — в состоянии покоя.
Оболочки камбиальных клеток иногда различаются не только по толщине, но и по своим коллоидным свойствам. Во время периода покоя стенки камбиальных клеток бывают наиболее толстыми и имеют свойства твердого геля, а во время активной деятельности оболочки становятся тоньше и более гибкими. Те специфические изменения, которые испытывают оболочки камбиальных клеток при дифференциации камбия в элементы ксилемы и флоэмы, заметно варьируют в зависимости от типа образующейся ткани. При формировании паренхимных элементов флоэмы, сохраняющих свою способность к росту и вытягиванию, исходные камбиальные клетки лишь немного видоизменяются в своей форме и по толщине оболочки, вторичных же утолщений оболочки почти не образуется. Наоборот, в камбиальных клетках, подвергающихся необратимому изменению, вследствие которого образуются слои вторичных утолщений оболочек, первичные оболочки во время процесса вытягивания клетки перед началом ее дифференциации становятся тоньше. Оболочка камбиальной клетки растягивается по мере того, как клетка увеличивается в размерах, но слои вторичного утолщения не откладываются до тех пор, пока клетка не достигнет окончательного размера.