Еще в верхушке побега под влиянием образования листьев и почек формируется его сложная проводящая система как единое целое.
Когда изучается следование проводящих пучков в стебле взрослого растения, необходимо иметь в виду, что их слияние и расщепление произошли еще в меристематическом состоянии, в период заложения органов.
Совокупность тканей стебля, расположенных внутрь от коры, называется центральным цилиндром, или стелой. Центральный цилиндр на ранних стадиях своего образования представляет собой в основном производное васкулярной меристемы — прокамбия. Снаружи он граничит с первичной корой (до тех пор, пока она существует), а внутри — с сердцевиной. Наружной тканью стелы является перицикл, состоящий из одного или нескольких слоев клеток. В стеблях перицикл часто бывает однослойным. Перицикл представляет собой образовательную ткань и способен к увеличению числа слоев составляющих его клеток.
Из перицикла образуются первичные лубяные волокна (вторичные — из камбия), в корнях он является корнеродным слоем — в нем зарождаются боковые корни. Кроме лубяных волокон, в перицикле залагаются глубокие слои перидермы, а также формируются млечные трубки и другие вместилища продуктов секреторной деятельности растения.
Вообще образовательная деятельность перицикла весьма разнообразна, поэтому его еще называют перикамбием. В старых корнях клетки перицикла одревесневают и даже опробковевают, но сравнительно редко.
Перицикл является пограничной тканью, заключая основные ткани центрального цилиндра — флоэму и ксилему. Выше уже отмечалось, что в зависимости от происхождения (из прокамбия или из камбия) элементы флоэмы и ксилемы обозначают как первичные и вторичные. Самые ранние элементы первичной ксилемы и первичной флоэмы называют протоксилемой и протофлоэмой.
У некоторых травянистых растений нередко вся ксилема является «первичной». Первичная ксилема с внутренней стороны часто облитерируется и разрушается, но наиболее поздно возникшие сосуды этой первичной ксилемы у большинства травянистых растений сохраняются. В многолетних побегах у древесных растений первичная ксилема сохраняется редко.
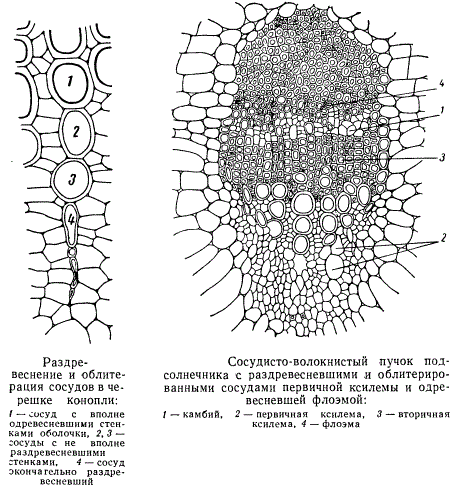
Протоксилема в стеблях никогда не сохраняется. Только в гипокотиле (подсемядольном колене) иногда заметны ее остатки, да и то на сравнительно молодой стадии развития гипокотиля. В корнях протоксилема сохраняется чаще.
В процессе развития растения ряд структурных изменений происходит также и во флоэме. Ситовидные трубки первичной флоэмы нередко облитерируются и разрушаются подобно сосудам первичной ксилемы. Флоэма способна одревесневать, в особенности у растений, образующих корку. Одревеснение флоэмы у травянистых растений происходит под осень и, по-видимому, связано со старением листьев. У большинства растений флоэма и ксилема объединены в пучки, которые сливаются в стеле в оплошное кольцо, не теряя, однако, в большинстве случаев полностью своей индивидуальности. В своем развитии сосудисто-волокнистые пучки стебля связаны с развитием определенных листьев. Поэтому сосудисто-волокнистые пучки стебля называют листовыми следами.
Исследования показали, что развитие проводящей системы стебля тесно зависит от развития листьев. Если отрезать какой-либо начинающий развиваться лист (листовой зачаток) и спустя некоторое время после операции сделать поперечный разрез стебля ниже места прикрепления ранее срезанного зачатка листа, можно убедиться, что сосудисто-волокнистые пучин, связанные с жилками отрезанного молодого листа, имеют ненормальный вид или совсем не развиваются. Степень дегенерации проводящего пучка стебля зависит от того, связан ли пучок с одним отрезанным развивающимся листом или при посредстве анастомозов соединен с пучками, идущими от других листьев, а также от того, на какой стадии развития был отрезан начинающий развиваться лист.
Под воздействием развивающегося листа участки васкулярной меристемы (прокамбия), расположенные в стебле ниже места прикрепления листа, постепенно превращаются в элементы сосудисто-волокнистого пучка. Новые сосудисто-волокнистые пучки, дифференцирующиеся из межпучкового камбия в сформировавшихся уже участках стебля, представляют собой нижние концы листовых следов, возникших при образовании новых листьев, зачатки которых появились на конусе нарастания стебля при дальнейшем развитии стебля. Сосудистые пучки как бы проникают из листа в стебель. Дифференцируясь дальше, звено за звеном вниз по стеблю, вновь образовавшиеся листовые следы встречают на своем пути более старые листовые следы, идущие от нижерасположенных листьев, и сливаются с ними. Таким путем создается сеть сосудистых пучков стебля.
Вполне сформировавшиеся «сосудисто-волокнистые пучки проходят вдоль по стеблю по нескольку междоузлий в зависимости от расположения листьев на стебле, а потом сливаются с расположенными рядом пучками. У подсолнечника сосудисто-волокнистый пучок как чистый листовой след, не сливаясь с соседними пучками, проходит от 2 до 5 междоузлий. До тех пор, пока сосудисто-волокнистый пучок, следуя вниз по стеблю, не сливается с другими пучками, его можно назвать индивидуальным листовым следом. Когда же индивидуальный листовой след соприкасается с другими листовыми следами и сливается с ними, образуется синтетический листовой след. В синтетическом листовом следе, на некотором расстоянии от места слияния, еще различимы отдельные индивидуальные листовые следы, образовавшие синтетический листовой след. Позже, особенно после причленения еще новых листовых «следов, границы между ранее слившимися листовыми следами совершенно исчезают.
Листовые следы стебля подсолнечника в верхней и средней зонах стебля на значительном расстоянии строго индивидуальны. Только в самом низу, начиная со второго яруса расположения листьев и ниже, листовые следы сливаются с соседними сравнительно скоро по внедрении. Индивидуальные листовые следы подсолнечника отграничены от соседних листовых следов мощными паренхимными прослойками (сердцевинными лучами) и не анастомозируют между собой.
В индивидуальном листовом следе, на пути его от места внедрения до места слияния, наблюдается значительное сокращение числа сосудов первичной ксилемы и одревеснение более старых участков флоэмы.
Размеры поперечника индивидуального листового следа по его длине меняются незначительно. Заметнее изменения в конфигурации очертаний поперечного среза.
Камбий в индивидуальном листовом следе функционирует очень слабо, но стоит такому листовому следу войти в тесное соприкосновение с другими листовыми следами, как происходит резкое изменение активности камбия, откладывающего здесь типичные элементы вторичной флоэмы и ксилемы.
Листовые следы переходят из листа в стебель под более или менее большим углом. На поперечных срезах через верхнюю зону узла можно видеть, что они расположены в коре стебля, но на более низких уровнях в зоне узла пучки отодвигаются от периферии, и, наконец, под узлом они занимают постоянное положение в центральном цилиндре вместе с другими пучками, связанными с вышерасположенными листьями. Этот процесс называется внедрением листовых следов в проводящий цилиндр стебля.
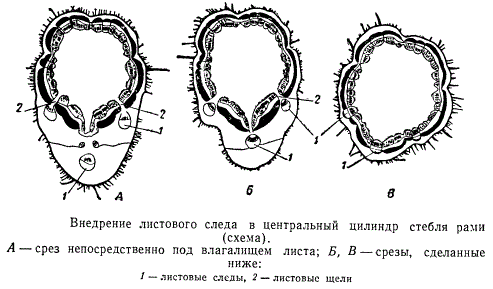
Следует указать, что выражение «листовой след внедряется» имеет лишь фигуральное значение, употребляемое для более наглядного определения структурного процесса, происходящего в центральном цилиндре. Пока листовой след не займет своего места в центральном цилиндре, против него в последнем остается более или менее широкий луч паренхимы, который называют листовой щелью, или листовым прорывом.
Ширина листовой щели, естественно, сильно зависит от размеров листового следа. От размеров же листового следа у травянистых растений непосредственно зависит и толщина массива ткани центрального цилиндра. Размеры самого листового следа в значительной мере обусловлены размерами пластинки листа. У льна — растения с относительно мелкими листьями — листовой след слабый, толщина ткани центрального цилиндра на поперечном разрезе стебля небольшая и листовая щель неширокая. У рами и подсолнечника листья широкие, соответственно листовые следы более мощные и листовые щели шире.
Более детально структурные изменения в центральном цилиндре стебля рами, связанные с внедрением листового следа, показаны на рисунке. На рисунке изображена периферическая часть поперечного разреза стебля рами, сделанного через середину междоузлия.

В середине междоузлия, где структура центрального цилиндра однородна, массивы лубяных волокон и ксилемы стебля располагаются у рами концентрическими почти непрерывными кольцами. Ткань твердого луба рами построена очень однообразно. Отдельные лубяные волокна или непосредственно соприкасаются друг с другом, или разделены между собой тонкостенной паренхимой. Элементы ксилемы по своему составу несколько разнообразнее элементов твердого луба, но вообще также однообразны. Широкие сосуды, сравнительно тонкостенные, вкраплены среди весьма толстостенных элементов либриформа и заменяющих волокон, причем последних значительно меньше, чем либриформа. Мало также и древесинной паренхимы.
По мере приближения к узлу стебля в секторе центрального цилиндра, расположенном непосредственно над местом прикрепления листа, постепенно проявляется ряд структурных изменений. В этом можно убедиться, делая идущие друг за другом сверху вниз последовательные поперечные срезы стебля.
На рисунке, изображающем срез, сделанный над узлом в непосредственной близости к нему, видно, что здесь как бы намечается структурная подготовка к формированию листовой щели.

Прежде всего массивы лубяных волокон разъединены, промежутки между ними заполнены тонкостенной паренхимой. В ксилеме, в участке, расположенном против участка паренхимного промежутка в твердом лубе, также заметны изменения. Толстостенные элементы ксилемы заменились относительно менее толстостенными элементами, напоминающими в своей совокупности ткань сердцевинных лучей. Особое внимание обращает на себя отсутствие первичной ксилемы. Делая последовательные срезы, можно удостовериться, что первичная ксилема исчезает над щелью несколько раньше, чем проявляются прочие структурные изменения центрального цилиндра в области узла.
При дальнейшем приближении к месту, занимаемому листовой щелью, вслед за исчезновением сосудов первичной ксилемы становится все меньше и меньше широкополостных сосудов во вторичной ксилеме и толстостенные элементы вторичной ксилемы все больше заменяются элементами менее толстостенными. Прозенхимные анатомические элементы ксилемы заменяются типично паренхимными, т. е. короткими клетками: на поперечном разрезе через эти участки узла почти каждая клетка имеет оболочки с порами. Все поры в паренхимных клетках простые.
В тонкостенной паренхиме, разделяющей массивы твердого луба, появляются друзы оксалата кальция. Дальнейшая фаза структурного метаморфоза тканей центрального цилиндра при образовании листовой щели характеризуется полной паренхиматизацией соответствующих участков. Строение листовой щели в верхней и нижней частях стебля травянистого растения несколько различно. В верхних частях стебля листовые щели шире, чем в нижних.

Если в пазухе листа находится ветка и она сравнительно далеко отстоит от места прикрепления листа, то щель сначала расширяется для внедрения сосудистых пучков, идущих от ветки в стебель, затем несколько суживается и опять расширяется для сосудистых пучков, идущих от листа. Первое расширение щели называется щелью ветки, в отличие от листовой щели.
Число листовых следов, идущих от каждого листа в центральный цилиндр, может быть различно. Как указывалось выше, у рами и подсолнечника от каждого листа в центральный цилиндр внедряется по три листовых следа; у льна, а также у ясеня только одна жилка листа продолжается в листовой след; у тополя и клена в каждый лист входят три, а у винограда — семь листовых следов. У многих зонтичных и лютиковых от каждого листа внедряется по нескольку листовых следов.
В соответствии с различным числом листовых следов неодинаково число образующихся при их внедрении в центральный цилиндр листовых щелей. Обычно для каждого листового следа формируется отдельная листовая щель. Но бывает также, что несколько листовых следов внедряются через одну щель. Вариации в количестве листовых следов и щелей присущи определенным систематическим группам растений, поэтому изучение строения узлов у различных растений, сопоставление особенностей внедрения листовых следов в центральный цилиндр в области узла имеет значение при исследованиях в области филогении.
Строение стелы в зависимости от характера ветвления и расположения почек и листьев отличается большим разнообразием. Например, организация стелы однодольных растений резко отличается от двудольных. У однодольных в зачатках листьев уже на ранних этапах их развития залагается большое количество прокамбиальных, а затем проводящих пучков. Основания листьев у однодольных кольцом охватывают стебель, их многочисленные проводящие пучки не умещаются в одно кольцо, как у двудольных, пучки же в стебле дифференцируются по всей толще меристемы. Так, у кукурузы, отличающейся большим количеством пучков, они расположены в несколько циклов до самого центра стебля, поэтому и сердцевины как структурно обособленной ткани у кукурузы нет. У пшеницы и риса с менее мощными листьями сосудистые пучки умещаются в 2—3 цикла, сердцевина же в процессе дальнейшего роста стебля разрушается. Сосудисто-волокнистые пучки однодольных растений в стебле идут всегда раздельно, не сливаясь от узла до узла. Только в узлах образуются сплетения сосудисто-волокнистых пучков вследствие развития густой сети анастомозов. Но настоящего слияния, как у двудольных, у однодольных не происходит. Камбия в сосудисто-волокнистых пучках однодольных нет или время деятельности его очень кратковременно и ограничено лишь первыми фазами развития вторичной ксилемы. Сосудисто-волокнистый пучок однодольного является истинным индивидуальным листовым следом.
Как видно, сосудисто-волокнистый пучок однодольного растения заключен в футляр из механических элементов, как бы изолирующих его от соседних тканей. Сосудисто-волокнистые пучки двудольных окружены только сравнительно тонкостенной паренхимой.