Одной из причин, изменяющих расщепление в отношении 3:1, является разная жизнеспособность зигот F2.
Примерами этого могут служить факты, обнаруженные при изучении наследования окраски у желтых и черных мышей, серой (ширази) и черной (араби) окраски у каракульских овец, платиновой и Серебристо-черной окраски чернобурых лисиц, наличия и отсутствия чешуи у зеркального карпа, ряда признаков у плодовой мушки дрозофилы и других животных, а также у растений и микроорганизмов. Рассмотрим некоторые из этих примеров.
Еще в начале нашего столетия было замечено, что при скрещивании желтых мышей между собой в потомстве наблюдается расщепление по окраске на две желтые мыши и одну черную. Позднее такое же явление было найдено у ряда других животных.
В 30-х годах была обнаружена новая окраска у чернобурой лисицы, названная платиновой. При скрещивании платиновых лисиц между собой появлялись как платиновые, так и серебристочерные лисицы в отношении соответственно 2:1. Природа этого явления была вскоре разгадана. Если форму с платиновой окраской представить как доминантную гомозиготу, т. е. АА, а серебристочерную — как рецессивную гомозиготу аа, то гибридное потомство должно быть, однообразным Аа и платиновой окраски. Однако этого не наблюдалось: потомство расщеплялось по окраске в отношении 1:1, т. е. напоминало расщепление при анализирующем скрещивании: АаХаа = 2Аа : 2аа, или 1Аа : 1аа. Когда же скрещивали лисицу платиновой окраски с таким же самцом, то имело место расщепление в необычном отношении 2:1. Отсюда стало очевидным, что платиновая окраска является доминантной, но пришлось предположить, что определяющий ее фактор находится в гетерозиготном состоянии. Тогда при скрещивании гетерозиготных лисиц Аа в потомстве должно было бы получаться менделевское расщепление 1АА : 2Аа : 1аа, а получалось 2Аа : 1аа, и гомозиготные платиновые лисицы АА никогда не обнаруживались. Поэтому предположили, что они не выживают и гибнут на эмбриональной стадии. Вскрытие беременных самок подтвердило это.
Следовательно, наследственный фактор, определяющий платиновую окраску лисиц, является доминантным, но связан с каким-то наследственным дефектом в развитии организма. Особи, у которых этот ген находится в гомозиготном состоянии, не выживают. Это заключение вытекает из того, что лишь гетерозиготные платиновые лисицы нормально развиваются и плодятся. Таким образом, звероводческие хозяйства, не знающие характера наследования этого признака и пытающиеся разводить платиновых лисиц «в себе», т. е. скрещивая их между собой, теряют 25% приплода в каждом поколении.
Такой же тип наследования был обнаружен при разведении овец, дающих серый каракуль, и в этом случае при скрещивании внутри себя наблюдается расщепление 2:1. Ягнята, гомозиготные по доминантной аллели гена, определяющего окраску ширази, гибнут после рождения от недоразвития желудочно-кишечного тракта.
Сходный пример можно привести из опытов В. С. Кирпичникова на зеркальных карпах. Среди культурных карпов имеются формы, четко отличающиеся по чешуйчатому покрову, так называемые чешуйчатые карпы (разбросанные, линейные) и голые. Эти различия обусловлены наследственно и связаны с некоторыми другими признаками: у линейных и голых карпов наблюдается сильная редукция ряда органов, что вызывает снижение жизнеспособности и скорости роста. Следовательно, гены, определяющие развитие чешуи (у линейных и голых форм), связаны, подобно факторам платиновой окраски лисиц, с наследственным дефектом в развитии организма, вызывающим летальный (смертельный) исход в гомозиготном состоянии.
Действительно, при скрещивании двух линейных карпов в потомстве наблюдается расщепление по характеру чешуи в отношении 2 линейных : 1 чешуйчатый. Если формы, гомозиготные по линейному расположению чешуи А, гибнут на эмбриональной стадии, то скрещивание можно представить так, как это изображено на рисунке.
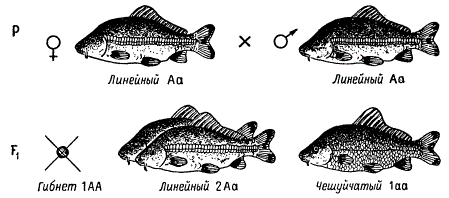
Наследование характера чешуйчатого покрова Cyprinus Carpio
Для осуществления нормального расщепления в F2 при моногибридном скрещивании необходимо, чтобы все зиготы, независимо от генотипа, имели равную жизнеспособность. Если это условие не соблюдается, то нормальное расщепление при моногибридном скрещивании в отношении 3 : 1 по фенотипу и 1 : 2 : 1 по генотипу может значительно видоизменяться.
Итак, уклонения от классического типа расщепления в отношении 3 : 1 ни в коей мере не могут поколебать закономерностей, установленных Менделем. Они лишь указывают на вмешательство дополнительных факторов и подчеркивают важность самого метода генетического анализа для изучения наследования признаков. Отношения при расщеплении по фенотипу и генотипу носят статистический характер. Именно поэтому Мендель использовал метод количественного анализа расщепления в потомстве по каждому из признаков родителей.
Итак, почему же в действительности может и не быть точного арифметического соответствия между классами расщепления по фенотипу и генотипу, хотя механизм мейоза обеспечивает равенство гамет с аллелями А и а у гибрида Аа? Объясняется это следующими обстоятельствами.
Как мы уже видели, анализ расщепления производится на основе учета признаков организмов, а не гамет, развивающихся у гибридов. Между образованием гамет и проявлением признаков организмов протекают сложнейшие процессы индивидуального развития. Для точного соответствия расщепления гамет и зигот необходимо:
- равновероятное расхождение хромосом в редукционном делении при образовании гамет; как мы увидим позднее, само расхождение хромосом в редукционном делении может нарушаться и приводить к неравенству образования двух типов гамет (А и а);
- равновероятное соединение всех сортов гамет при оплодотворении;
- равная жизнеспособность зигот с различными генотипами: АА, Аа и аа. Но мы уже видели из приведенных примеров, что гомозиготные формы по доминантным генам иногда оказываются нежизнеспособными, и целый класс расщепления выпадает, давая отношение 2 : 1 вместо 3 : 1. Кроме того, три генотипических класса зигот (АА, Аа и аа) могут отличаться не сильными, а слабо заметными изменениями жизнеспособности, вследствие чего также изменяется численное соотношение классов;
- полное проявление признака независимо от изменения его под влиянием факторов внешней среды (так, например, если подсчитывать расщепление среди семян гороха по окраске — желтой или зеленой, то недозрелые семена будут давать промежуточное выражение признака, что затрудняет объективную классификацию их).
Все перечисленные моменты, а также малое количество особей в потомстве гибрида (семьи) могут давать отклонения от точного соотношения классов расщепления. В основе расщепления по фенотипу и генотипу действительно лежит статистическая закономерность. Но для проявления всякого закона требуются условия, при которых он может осуществляться. Расщепление в потомстве гибрида является биологическим законом, осуществляющимся с математической точностью при образовании гамет. Подсчет же фенотипов производится при анализе организмов, и характер расщепления, следовательно, устанавливается на основе экстраполяции от признака к гену. Чтобы показать справедливость сказанного, обратимся к анализу расщепления гамет.