По поводу механизма перекреста существует несколько гипотез, но ни одна полностью не объясняет установленных фактов рекомбинации генов и наблюдаемые при этом цитологические картины.
Как мы уже указывали, на первом этапе изучения кроссинговера его связывали непосредственно с образованием хиазм. Но при этом оставалось неясным, являются ли хиазмы следствием произошедшего кроссинговера или они предшествуют ему и необходимы для его осуществления. Альтернатива этих событий сводится к двум противоположным возможностям: 1) вначале образуются хиазмы, затем происходит разрыв и обмен гомологичными участками, и 2) сначала происходит разрыв, а затем обмен и образование хиазм. Различия этих возможностей показаны на рисунке в виде двух схем.
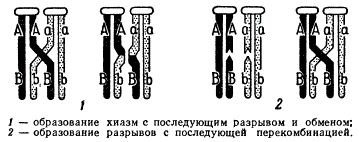
Схемы, иллюстрирующие различные гипотезы механизма перекреста хромосом
Согласно гипотезе Ф. Янсенса, предложенной в 1909 г. и развитой К. Дарлингтоном в 1937 г., в процессе синапсиса гомологичных хромосом и образования бивалента в последнем создается динамическое напряжение, которое складывается из ряда взаимодействующих сил, возникающих в связи со спирализацией хромосомных нитей, а также при взаимном обвивании гомологов в биваленте. В силу возникающего динамического напряжения одна из четырех хроматид рвется. Разрыв нарушает равновесие в биваленте. По оси гомологичного партнера создается избыточное напряжение, которое приводит к компенсирующему разрыву в строго идентичной точке одной из его хроматид. Напряжение в биваленте падает и начинается раскручивание гомологов — происходит терминализация хиазм, в ходе которой возможно реципрокное воссоединение разорванных концов, приводящее к кроссинговеру. Согласно этой гипотезе хиазмы непосредственно связаны с кроссинговером.
По гипотезе К. Сакса, предложенной в 1930—1932 гг., хиазмы не являются результатом кроссинговера: сначала образуются хиазмы, а затем происходит обмен. При расхождении хромосом к полюсам вследствие механического напряжения в местах хиазм происходят разрывы и обмен соответствующими участками. После обмена хиазма исчезает. Согласно другой вариации этой гипотезы (X. Мацуура), между образованием хиазм и кроссинговером нет никакой связи. Кроссинговер осуществляется при синапсисе несестринских хроматид и расхождения их в анафазе I, когда центромеры расходятся к полюсам. Как видно, обе гипотезы исходят из механических причин кроссинговера.
Гипотеза Дж. Беллинга (1933 г.) сводится к предположению, что кроссинговер приурочен к репродукции хромосом и не связан с предварительным их разрывом. В первоначальном виде, исходя из корпускулярного строения хромосом, Беллинг представлял, что отдельные хромомеры, содержащие гены, в процессе репродукции хромосом могут объединяться в новые сочетания. Такое объединение репродуцированных хромосом и обеспечивает кроссинговер.
Позднее (1955 г.) эта гипотеза была модернизирована И. Ледербергом для объяснения рекомбинации у микроорганизмов. Смысл данной вариации заключается в том, что в процессе репликации воспроизведение последней может реципрокно переключаться с одной нити на другую: воспроизведение, начавшись на одной матрице, с какой-то точки переключается на матричную нить ДНК гомологичного партнера, а затем вновь возвращается на свою матрицу, что и приводит к рекомбинации генетического материала. Эта гипотеза была названа гипотезой «выбора копии» (сору choice).
В 1963 г. была выдвинута еще одна гипотеза кроссинговера, автор X. Уайтхауз допускает возможность дополнительной репликации гибридной молекулы ДНК в процессе рекомбинации. Эта гипотеза пытается объяснить кроссинговер с точки зрения молекулярных процессов. Она может быть пригодной для ряда случаев рекомбинации. Однако допущение дополнительной репликации ДНК в процессе рекомбинации на хроматидном уровне делает эту гипотезу маловероятной.
Таким образом, ни одна из гипотез пока не объясняет полностью процесса кроссинговера. Основной трудностью является то, что репликация молекулы ДНК происходит в интерфазном состоянии ядра, а для рекомбинации необходимо осуществление синапсиса гомологичных хромосом в профазе I, если кроссинговер действительно происходит на хроматидном уровне. Если принять молекулярный механизм кроссинговера, то следует признать, что профаза I вообще не имеет отношения к кроссинговеру, и последний осуществляется в интерфазном состоянии ядра, синапсис же хромосом имеет какое-то другое назначение.