Для того чтобы произошел перекрест гомологичных хромосом, необходимо предварительное попарное их соединение — синапсис.
В норме это происходит только в профазе I.
Причины и механизм синапсиса остаются невыясненными. Предполагают, что гены, находящиеся в идентичных локусах хромосом, приобретают свойство взаимного притяжения как разноименно заряженные частицы, или что они выделяют при этом особые вещества, обеспечивающие, своеобразный положительный «таксис» хромосом.
Хромосомные нити каждой из гомологичных хромосом на стадии профазы I имеют определенную локализацию хромомер, которые являются своего рода морфологическими маркерами каждой пары гомологичных хромосом: в идентичных участках хромосом они оказываются сходными по величине и морфологии. Нельзя представлять, что хромомеры аналогичны генам, но их постоянство по форме их числу в каждой хромосоме дает основание считать, что образование их наследственно детерминировано. По ним можно узнать любую хромосом в наборе каждого вида на лептотенной стадии профазы I. В зиготенной стадии сходные хромомеры притягиваются друг к другу и располагаются попарно. В результате синапсиса образуются биваленты. В следующей, диплотенной стадии происходит перекручивание хроматид, в результате которого обращаются хиазмы. На этой же стадии центромеры начинают делиться и расходиться, и теперь только хиазмы удерживают хроматиды бивалента в одном комплексе.
По мере расхождения центромер к противоположным полюсам происходит терминализация хиазм. В силу терминализации хиазм и продолжающейся спирализации хроматид образуются различные фигуры бивалентов, или тетрад (стадия диакинеза).
В зависимости от того, произошел ли перекрест и если произошел, то был ли он одинарным или двойным, фигуры бивалентов изменяют свою форму.
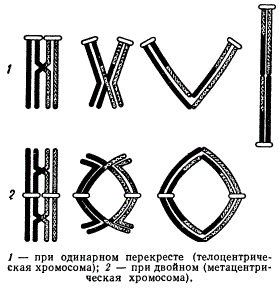
Схема терминализации хиазм
При одинарном перекресте в телоцентрических хромосомах (т. е. когда центромера находится на конце хромосомы) хиазмы сползают от центромеры к дистальным концам хромосом, и в диакинезе образуются палочковидные фигуры бивалента. При двойном перекресте в метацентрической хромосоме (т. е. когда центромера расположена в середине хромосомы) биваленты образуют кольца. Иногда эти фигуры могут быть и более сложными.
На первом этапе изучения кроссинговера механизм его казался довольно простым. Сначала полагали, что рекомбинация генов происходит в силу разрывов в идентичных участках гомологичных хромосом и последующего их обмена равными сегментами. Момент кроссинговера приурочивали к стадии профазы I, начиная с синапсиса зиготенной стадии, и считали, что он происходит между гомологичными одиночными хромосомными нитями или уже удвоенными хромосомами. Последнее вытекало из того, что в мейозе двум делениям ядра соответствует одна репродукция хромосом. Теперь установлено, что репликация ДНК происходит в интерфазе. Следовательно, хромосомы уже в лептотенной стадии являются двойными.
На основании генетического анализа твердо установлено, что кроссинговер происходит на стадии четырех хроматид. Доказательства этому были получены на дрозофиле с помощью различных моделей скрещивания при соматическом кроссинговере, на трисомиках кукурузы, а также на нейроспоре и дрожжах с помощью тетрадного анализа. Не вызывает сомнений также то, что кроссинговер приурочен к образованию хиазм, хотя причинно-следственная связь этого пока остается необъясненной.
Частота мейозов, во время которых происходит обмен идентичными участками между гомологичными хромосомами, зависит от расстояния между изучаемыми генами. Чем дальше отстоят друг от друга гены, тем более возможен обмен. Однако процент кроссоверов к общему числу учитываемого потомства никогда не превышает 50%. Допустим, что произошел одинарный кроссинговер, т е. в одном биваленте образовалась одна хиазма, и из четырех хроматид бивалента обменялись идентичными участками только две несестринские хроматиды.
Две другие несестринские хроматиды каждой хромосомы остаются не затронутыми кроссинговером. Очевидно, что в каждом биваленте 50% хроматид, или будущих хромосом, оказываются рекомбинантными и 50% — нерекомбинантными. В результате двух мейотических делений образуются две гаметы с родительским типом расположения генов и две — с рекомбинантным.
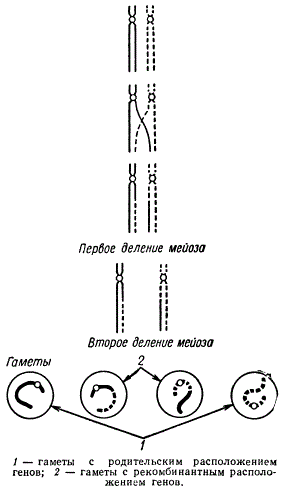
Схема образования гамет после одинарного кроссинговера
Следовательно, если даже во всех клетках (т. е. 100%), претерпевших мейоз, произошел одинарный кроссинговер, то и в этом случае только половина гамет (50%) будет нести кроссоверные хромосомы.
Но чем больше расстояние между генами, тем чаще в мейозе будут встречаться обмены, и наоборот. Например, если из 1000 клеток, в которых прошли оба деления мейоза, одинарный кроссинговер осуществился только в 100 случаях, то из 1000⋅4 образующихся гамет 200 окажутся рекомбинантными, и 200 — нерекомбинантными. Средний процент рекомбинаций на все учтенные гаметы составит 200/4000⋅100 = 5%.
Этот процент указывает на частоту кроссинговера между двумя учитываемыми генами. Если же кроссинговер произойдет не в 100, а в 200 клетках, вступивших в мейоз, из 1000, то исходя из частоты кроссинговера расстояние между генами должно быть в два раза больше 400/4000⋅100 = 10%.
Теперь предположим, что в одном биваленте прошел не один кроссинговер, а два и более. При этом каждая хроматида хромосомы бивалента может обмениваться независимо в любом сочетании с хроматидами другой хромосомы того же бивалента, и тогда образуется несколько хиазм. Поскольку в биваленте четыре хроматиды, то очевидно, что каждая из них имеет равную вероятность обменяться участками с любой другой. При этом в обмене могут участвовать две, три или все четыре хроматиды, но в любом месте каждая хроматида вступает в обмен с несестринской хроматидой другой хромосомы только один раз.
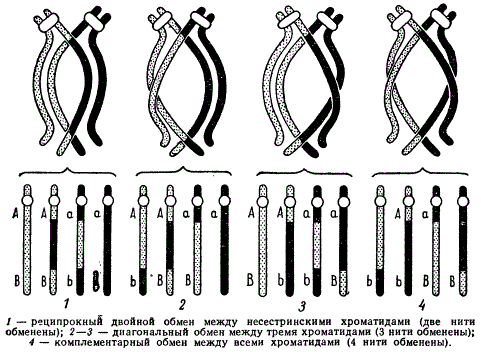
Схема двойных хиазм и продукты двойных обменов между хроматидами тетрады
Приведена схема подобных обменов:
1) реципрокного двойного обмена между двумя несестринскими хроматидами, не дающего рекомбинаций генов, если гены-маркеры не затронуты обменом;
2) диагонального обмена, когда две сестринские хроматиды в двух разных районах одновременно вступают в одинарный перекрест с одной и той же несестринской хроматидой, а четвертая хроматида не вовлечена в обмен (в результате такого двойного обмена возникают три рекомбинантных хромосомы и одна остается нерекомбинантной);
3) комплементарного обмена, когда все четыре хроматиды претерпевают одинарные обмены в разных районах, две несестринские хроматиды из четырех попарно претерпевают одинарный обмен в одном месте, а две другие — во втором, вследствие чего возникают четыре рекомбинантных хромосомы.
Двойные кроссоверы могут возникать как следствие одновременных одинарных обменов между хроматидами при участии в обмене трех хроматид бивалента. Если учесть суммарно все комбинации хроматид при двойном обмене: некроссоверные хроматиды, хроматиды с одинарным и хроматиды с двойным обменом, то получим отношение 4:8:4, или 1 : 2 : 1, где первый класс — некроссоверные хроматиды, второй — одинарные и третий — двойные кроссоверные хроматиды. Специальные исследования показали, что у ряда организмов перекрест в одном месте между двумя любыми сестринскими хроматидами не влияет на перекрест между двумя любыми хроматидами в других местах того же бивалента. Это доказывает, что в данном случае хроматидная интерференция отсутствует: перекрест между одними хроматидами не влияет на перекрест между другими хроматидами.
В том случае, когда гены-маркеры не вовлекаются в двойной кроссинговер, рекомбинантные хроматиды не будут обнаружены, и проявятся только одинарные. Если данная пара хромосом в биваленте была бы маркирована тремя парами генов в гетерозиготном состоянии ABC/abc то для пары генов А С, расположенных вне обменного участка, отношение некроссоверных к кроссоверным составит 2 : 2.
В приведенных выше расчетах мы исходили из того, что перекрест хроматид в гомологичных хромосомах осуществляется случайно и каждый обмен не влияет на другой. Однако было указано на существование явления интерференции, когда нарушается случайность обмена. Тогда множественные обмены Могут возникать либо реже (положительная интерференция), т. е. один обмен подавляет другой, либо чаще (отрицательная интерференция), т. е. один обмен способствует другому, в сравнении с частота при отсутствии их взаимного влияния.
Различают хроматидную интерференцию и хиазменную интерференцию. Под хроматидной интерференцией имеют в виду отклонение от случайного участия в обмене всех четырех хроматид бивалента т. е. одни хроматиды вступают в обмен либо чаще, либо реже, другие. В этом случае наблюдаются отклонения результатов разных типов двойных обменов от отношения 4 : 8 : 4 (или 1 : 2 : 1). Хроматидная интерференция может быть положительной и отрицательной. Положительная хроматидная интерференция имеет место в том случае, когда участие одной и той же хроматиды в одном обмене препятствует ее участию во втором обмене, отрицательная — тогда, когда одна и та же хроматида предпочтительно вовлекается оба обмена.
Хиазменной интерференцией называют положительное или отрицательное взаимовлияние обменов друг на друга по длине бивалента независимо от участвующих хроматид. Оба типа интерференции зависят от структуры хромосом, места обмена и других факторов. Отрицательная интерференция может встречаться в бивалентах различных организмов, но практически выявляется только тетрадном анализе, где непосредственно контролируются проекты мейоза.
Как мы указывали, между кроссинговером и образованием хиазм предполагается прямая связь, хиазмы являются внешним проявлением идущего или прошедшего обмена участками между хроматидами гомологичных хромосом. Поскольку кроссинговер может быть множественным, то очевидно, что и число хиазм может быть более одной на бивалент. Так как каждая хиазма образуется за счет обмена между каждыми двумя несестринскими хроматидами из четырех, то число обнаруживаемых клеток (мейозов) с хиазмами оказывается всегда в два раза больше, чем частота обнаруживаемого кроссинговера.
Если между числом хиазм и частотой кроссинговера существует соответствие, то его можно проверить. Для этого К. Дарлингтон провел оценку предельных размеров длины генетических карт хромосом кукурузы на основе подсчета числа хиазм в профазе I. Он установил, что на 10 бивалентов в мейозе (клеток) у кукурузы приходится в среднем 27 хиазм. Так как биваленты имеют различную длину, то он внес поправку на длину каждого бивалента. Дарлингтон рассчитал, что 1 хиазма соответствует расстоянию в 50 морганид на генетической карте. 2,0 хиазмы на один бивалент соответствуют расстоянию в 100 морганид, 3,5 хиазмы — в 175 морганид и т. д.
Несмотря на то, что число локализованных генов в группах сцепления кукурузы за 16 лет изучения значительно возросло и соответственно генетические карты хромосом «удлинились», тем не менее ни в одном случае не установлено превышения расчетной длины. По мере того как будут локализованы новые мутантные гены у кукурузы, расчетные данные станут более соответствовать эмпирическим.
Хиазмы чаще всего в норме наблюдаются в проксимальных участках хромосом. Параллельно с этим установлено, что кроссинговер также чаще осуществляется в проксимальных и реже в дистальных участках хромосом. Важно отметить, что при отсутствии хиазм кроссинговер, как правило, не происходит.
Существует определенная зависимость между числом хиазм и длиной хромосомы. Очень короткие хромосомы имеют лишь одну хиазму на один бивалент. С увеличением длины хромосом число хиазм увеличивается до 10. Но прямой зависимости между длиной хромосом и числом хиазм на бивалент все же не установлено.
До сих пор мы рассматривали кроссинговер между несестринскими хроматидами. Естествен вопрос, может ли происходить обмен участками, т. е. кроссинговер, между сестринскими хроматидами, которые в профазе I мейоза и митоза длительное время находятся вместе. Такое допущение можно было сделать в отношении высших организмов, имеющих диплоидный набор хромосом и половое размножение. Оно очень вероятно для бактерий, где установлена кольцевая хромосома, как, например, у кишечной палочки.
Здесь мы не будем излагать всего хода экспериментального доказательства наличия кроссинговера между сестринскими хроматидами ввиду его сложности. Укажем только, что на специальных хромосомных моделях (кольцевых хромосомах) у кукурузы Б. Мак-Клинток и Д. Шварц в 1953 г. показали, что между сестринскими хроматидами также происходит обмен участками. Позднее с применением радиоавтографических методов было доказано наличие таких обменов у различных видов растений и животных и у человека.
Обмен внутри сестринских хроматид не может приводить к рекомбинациям, поскольку они генетически идентичны, и в силу этого он не имеет биологического смысла в качестве механизма наследственной комбинативной изменчивости.