Современные представления о формировании и развитии хлоропластов в онтогенезе растений возникли на базе электронно-микроскопических исследований. Наибольшее распространение получили две схемы развития ультраструктуры хлоропластов; одна из них была разработана Д. Веттштейном, а другая — К. Мюлеталером и А. Фрей-Висслингом.
Эти схемы в основном сходны, имеются лишь некоторые различия в деталях. Поэтому остановимся лишь на схеме Д. Веттштейна, который изучал развитие структурных элементов хлоропластов у разных видов растений с помощью электронного микроскопа и получил большое количество фотографий препаратов пластид ячменя, кукурузы, томатов, ели и других растений, выращенных в нормальных условиях освещения и в темноте, а также фотографии хлоропластов различных мутантов с нарушениями в пластидном аппарате. Поскольку никаких заметных различий между растениями разных видов обнаружено не было, Д. Веттштейн предложил общую схему развития ультраструктуры хлоропласта.
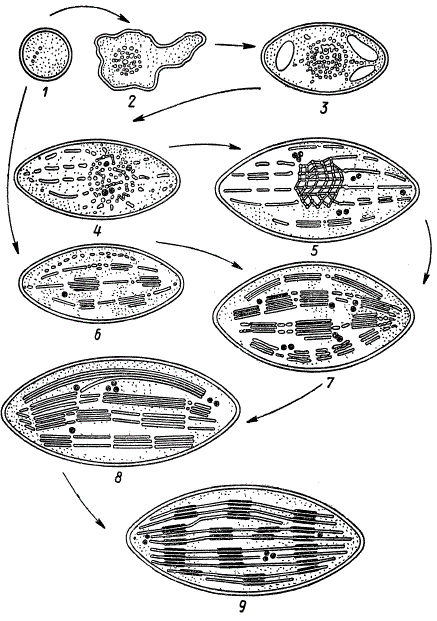
Схема развития хлоропласта (по Д. Веттштейну): этапы 1, 6—9 — нормальный ход развития на свету; этапы 1, 2 3, 4, 5 — путь развития пластиды в темноте
Пластиды, в частности хлоропласты, развиваются из пропластид, которые находятся в цитоплазме клеток меристемы листьев и точек роста. Эти предшественники пластид представляют собой мельчайшие органеллы округлой или овальной формы. Для пропластиды уже характерно наличие двойной мембраны, окружающей плотный матрикс, в котором имеется несколько пузырьков. Эти пузырьки либо попадают в пропластиду из окружающей цитоплазмы, либо формируются ее внутренней мембраной. При этом на внутренней мембране образуются впячивания внутрь матрикса, которые затем отшнуровываются от мембраны в виде пузырьков. По мере роста пропластиды увеличивается и число пузырьков в ней. Дальнейшее развитие зависит от условий произрастания растения. Если развитие идет на свету, то пузырьки по мере своего возникновения располагаются в несколько слоев. Вместе с тем они сливаются друг с другом в отдельные ламеллярные диски, которые также располагаются в несколько слоев. В строго определенных местах таких первичных слоев число дисков продолжает увеличиваться, в результате чего появляются пачки с 2—4 дисками примерно одинакового размера (этап 6). На последующих этапах развития возрастает число дисков, они становятся крупнее и сливаются друг с другом. Образовавшиеся слои состоят из плотно прилегающих друг к другу дисков; соединенных двойными ламеллами. Так, постепенно создается единая непрерывная ламеллярная система (этапы 7 и 8). После этого четко дифференцируются области, соответствующие гранам, благодаря строго упорядоченному расположению дисков (этап 9). На этом этапе полностью заканчивается дифференцировка структурных элементов хлоропласта.
Если же растение выращивается в темноте или при слабом диффузном освещении, то пузырьки не располагаются слоями и не сливаются в диски, а собираются в одном или нескольких местах, образуя плотные скопления, которые могут приобретать форму шариков. Эти скопления пузырьков получили название проламеллярных телец (этап 2), поскольку из них могут развиваться ламеллярные структуры хлоропласта при перенесении растения на хорошее освещение. Если же растение остается в темноте, то количество пузырьков продолжает увеличиваться, в результате чего проламеллярные тельца могут достигать настолько больших размеров, что становятся видимыми с помощью светового микроскопа. После того как в проламеллярном тельце соберется много пузырьков, начинается их слияние в сложную трубчатую структуру, сходную с кристаллической решеткой (этап 3). При дальнейшем увеличении размеров проламеллярного тельца некоторые пузырьки выходят из него и располагаются по радиусам (этап 4).
Если же растение перенести на хорошее освещение, проламеллярные тельца быстро перестраиваются, включаясь в ламеллярную систему хлоропласта. На рисунке это изображено схематически как переход от 5-го к 7-му этапу. Если же растение не подвергается воздействию света и остается в темноте, то дальнейшего развития пластид, достигших 5-го этапа, не происходит. Поэтому-то в них и не образуются ламеллярные структуры. Исключение из исследованных растений представляло развитие хлоропластов у ели.
Д. Веттштейн установил, что у проростков ели в отсутствие света наряду с синтезом хлорофилла происходит развитие нормальной ламеллярной структуры хлоропластов. Сам факт образования хлорофилла в темноте у проростков некоторых хвойных растений не нов, он был обнаружен еще до этих исследований. Здесь важно и интересно другое. По мнению Д. Веттштейна, для нормального размещения ламелл и создания определенной структуры хлоропласта необходимо наличие хлорофилла, хотя его присутствие не обязательно для возникновения пузырьков в пропластиде, а также, по-видимому, и для образования ламелл.
Согласно Д. Веттштейну, ламеллы в хлоропласте быстро обновляются благодаря распаду и восстановлению. Так, у ели накопление первичного крахмала содействует разрушению ламелл, а после удаления крахмала ламеллы образуются вновь.
Рассмотренная выше схема соответствует современным представлениям о развитии ультраструктуры хлоропластов, однако она охватывает только период от пропластиды до образования полностью сформированного хлоропласта. Данные А. А. Табенцкого, П. А. Генкеля и Р. С. Морозовой, а также других авторов показывают, что и дальнейшее развитие хлоропласта в онтогенезе растения, начиная с того момента, когда он полностью сформировался, сопровождается изменениями не только его размеров и формы, но и внутренней структуры.
Рассмотрим результаты электронно-микроскопических исследований А. А. Шахова и Б. М. Голубковой, свидетельствующие о больших изменениях структуры хлоропластов на протяжении вегетации растений картофеля. Эти исследования показали, что в ранние периоды вегетации можно наблюдать лишь незначительные различия между хлоропластами листьев, находящихся на разных ярусах одного и того же растения. Большие изменения в структуре хлоропластов происходят в более поздние периоды вегетации, и тогда обнаруживаются резкие различия между разновозрастными листьями. Так, у молодых растений (23-дневных, имеющих 8 ярусов) хлоропласты верхних, еще не закончивших рост листьев мало отличаются от хлоропластов нижних, календарно более старых листьев, но в обоих случаях они крупные (до 6—6,5 мкм) и округлые. Структурные их элементы четко дифференцированы: матрикс, хорошо очерченные крупные граны (0,5—0,7 мкм), располагающиеся довольно рыхло в матриксе, осмиофильные глобулы (мелкие, особенно у верхних листьев).
Хлоропласты листьев верхнего яруса остаются без заметных изменений и в более поздние периоды онтогенеза растений (фазы бутонизации и цветения). Им свойственна хорошо выраженная ламеллярно-гранулярная структура, как и листьям молодых (23-дневных) растений. Небольшое различие имеется в отношении осмиофильных глобул, которые становятся крупнее. В хлоропластах же листьев среднего и особенно нижнего ярусов этого же растения происходят значительные изменения структурных элементов. Размер хлоропластов уменьшается до 4,5—3 мкм, плотность увеличивается, граны некрупные и имеют тенденцию к исчезновению. Зато резко возрастает размер осмиофильных глобул до 0,3—0,45 мкм, а у нижних листьев — даже до 0,6 мкм. Образование осмиофильных глобул достигает максимума, и в конце концов они заполняют всю полость хлоропласта.
В результате дальнейшего старения наступает деструкция хлоропластов, когда на их месте образуется липоидная гранулярная масса, в которую включены крупные осмиофильные глобулы и крахмальные зерна.
Таким образом, эти исследования свидетельствуют о том, что в онтогенезе растений картофеля ярко выражены изменения структуры хлоропластов не только у листьев разных ярусов в пределах одного и того же растения, но и одних и тех же листьев в разные периоды вегетации.
Электронно-микроскопические исследования Т. Е. Кисляковой с соавторами показали, что листья средних ярусов в разные периоды онтогенеза растений картофеля имеют структурно наиболее организованные хлоропласты. Особенно высокая степень упорядоченности структуры характерна для фазы бутонизации: ламеллярная система их сильно развита, хорошо дифференцированные граны располагаются плотно и равномерно ориентированы по всему хлоропласту. Матрикс занимает главным образом постенный слой под оболочкой. Листья с такими хлоропластами отличаются наибольшей интенсивностью фотосинтеза. Последний вывод представляется особо важным и позволяет предположить, что структура хлоропластов листьев средних ярусов в фазу бутонизации картофеля является оптимальной для фотосинтетической деятельности.
Хорошо известно, что зеленый цвет листьев, а также незрелых плодов зависит от наличия в их клетках зеленых пластид, а желтая, оранжевая, красная и другая окраска многих плодов и лепестков цветков обусловлена присутствием окрашенных в соответствующие цвета пластид, общее название которых — хромопласты (от греч. хрома — цвет).
Кроме окрашенных, существуют и бесцветные пластиды — лейкопласты (от греч. лейкос — белый), характерные для неокрашенных частей растения, например для клеток корня. На протяжении вегетации пластиды одного типа могут превращаться в пластиды других типов. Так, при созревании плодов, осеннем изменении листьев и в других случаях хлоропласты переходят в хромопласты. Лейкопласты могут превращаться в хлоропласты при позеленении клубней картофеля или в хромопласты при формировании корнеплода моркови и т. д. Эти факты свидетельствуют о генетическом родстве разных типов пластид, благодаря которому и возможны такие их превращения.
Согласно А. Шимперу, пластиды не могут возникать заново, они образуются только путем деления уже существующих или в результате превращения пластид одного типа в пластиды других типов. Одна и та же пластида обладает способностью к определенным многократным изменениям, после чего она снова может вернуться в исходное состояние и приобрести прежние свойства.
Способность пластид одного типа превращаться в пластиды других типов А. Шимпер считал наиболее существенным их свойством. Схематически его представления о взаимопревращениях пластид можно изобразить следующим образом:

Стрелками на схеме указаны возможные направления превращений.
Дальнейшие наблюдения показали, что хлоропласты, потерявшие зеленую окраску и достаточно сильно пожелтевшие после перенесения растения в темноту, уже не способны снова стать зелеными и причиной этого является необратимая их деструкция. Равным образом и превращения хлоропластов в хромопласты оказываются необратимыми после полной утраты ими зеленой окраски. Отсюда некоторые исследователи делают вывод о том, что превращения пластид происходят не циклически, а лишь в определенных направлениях по схеме:

В процессе развития пластид лейкопласты непосредственно примыкают к пропластидам, что отражено на схеме и согласуется с действительностью, поскольку лейкопласты отличаются от пропластид лишь несколько большим размером.
Хромопласты образуются в результате накопления каротиноидов или из пропластид и лейкопластов, или из хлоропластов. Они возникают, как правило, из хлоропластов и значительно реже, как, например, в корнеплоде моркови, из лейкопластов. Хлоропласты возникают из пропластид либо из лейкопластов. Недавно были получены данные о переходе хромопластов в хлоропласты, а также о восстановлении нормальной структуры хлоропластов пожелтевших листьев под действием обработки кининами (стимуляторы роста). Эти данные говорят о возможности более широких взаимопревращений пластид, чем это представляется в соответствии с последней схемой.
Из всех пластид только хлоропласты имеют чрезвычайно тонкую и высоко упорядоченную структуру, без которой не происходят сложнейшие процессы поглощения и преобразования световой энергии в химическую энергию веществ, строящихся при фотосинтезе. Отсюда ясно, что переход хлоропластов в хромопласты должен сопровождаться разрушением их структуры. Действительно, образование хромопластов из зеленых пластид характеризуется большим или меньшим разрушением их ламеллярной структуры (иногда вплоть до полной деструкции), постепенным разрушением хлорофилла и накоплением каротиноидов, представляя собой, как считают многие ученые, конечный этап процесса развития и старения хлоропластов. Постепенное обесцвечивание зеленых пластид и превращение их в различно окрашенные хромопласты легко проследить путем наблюдений за развитием лепестков цветков у лютиковых и многих других растений или за созреванием плодов шиповника, рябины, перца и т. д.
Согласно А. Фрей-Висслингу, при распаде тонкой структуры зеленых пластид ценные для растения соединения, такие, как вещества, входящие в состав хлорофилла, а также другие азотсодержащие вещества, высвобождаются из структурных элементов хлоропластов и покидают стареющие ткани.
Вещества, менее ценные и легче доступные для растения, такие, как липиды и каротиноиды, накапливаются в хромопластах, которые, возможно, имеют значение резерва этих веществ.
Источник: Н.Н. Овчинников, Н.М. Шиханова. Фотосинтез. Пособие для учителей. Изд-во «Просвещение». Москва. 1972