Живые существа нашей планеты олицетворяют «уникальность биологического вещества» (В. И. Вернадский, 1967), т. е. имеют единую биохимическую организацию.
Есть веские доказательства в пользу того, что почти 93% всех ферментов, характерных для высших растений, играли роль в жизни древних прокариот (Э. Брода, 1978). Точно так же почти одинаковы механизмы фиксации CO2 и аэробного дыхания у зеленой водоросли и покрытосеменных, несмотря на значительный морфологический разрыв между ними. Сходны и механизмы поглощения веществ и испарения воды у наземных растений при огромном их многообразии. В связи с этим иногда говорят о прекращении эволюции веществ с наступлением эры биологической эволюции.
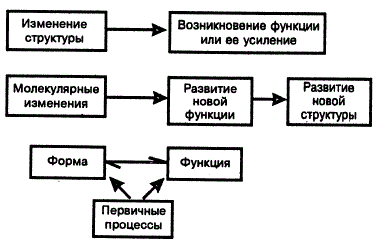
Здесь уместно остановиться на взаимосвязи формы (структуры) и функции. Этот вопрос является одним из древних и традиционных в натурфилософии и биологии. Долгое время он был оплотом идеалистических построений. В свое время Аристотель, задавая вопрос: «Ради чего существует орган?», отвечал: «Ради выполнения определенной цели», т. е. функции! Идеалисты исходили из тезиса, что животные и растения с известными органами и привычками якобы созданы для обитания в строго конкретных условиях.
Форма определяет функцию в онтогенезе, а функция — форму в филогенезе. Значение любой функции можно оценить объективно только в иерархии систем функций и структур. При наличии множества функций у структуры значимость последней возрастает при различных экологических ситуациях. Многие важные биологические функции обычно обеспечиваются несколькими структурами. Например, функция фотосинтеза протекает при участии листьев, стеблей, чашелистиков, плодов, остьев и т. п. Нередко монофункциональность на молекулярном уровне переходит в мультифункциональность на уровне организма. Так, функция молекулы хлорофилла на уровне организма расширяется: улавливание энергии света, фототаксис хлоропласта, дыхание и т. п.
Структуру (форма и орган) и функцию (физиологическое отправление) рассматривают в различных соотношениях, отдавая предпочтение то одной, то другой. Обычно первенство принадлежит изменению функции. Но это не всегда правильно. Вернее усматривать взаимосвязь и взаимообусловленность их изменений в эволюции. В «Происхождении видов» Ч. Дарвин писал, что «…для нас несущественно, изменяются ли сперва привычки, а затем строение органов…». Примеры, подтверждающие данное положение на животных, описал сам автор и другие исследователи.
Заметим, что все эти возможности встречаются и у растений; соответствующие примеры приведены ниже. Однако в ботанической литературе вопросу о взаимоотношении формы и функции уделяют мало внимания. Между тем растительное царство может дать не менее убедительные примеры взаимообусловленности изменения формы и функции в ходе эволюции. Выход растений на сушу, как известно, ознаменовался приобретением комплекса морфофизиологических новшеств: защитных покровов (эпидермы и кутикулы), проводящей системы, Дифференциации тела на органы и т. д. Подобными изменениями прежде всего было достигнуто уменьшение потери воды от испарения и усиление ее движения по растению. В приведенных примерах трудно сказать, что чему предшествовало, морфологические или физиологические изменения, хотя, несомненно, характер первых определял состояние организма в новых условиях. В то же время очевидно, что «заказ» на уменьшение отрицательных последствий недостатка воды повлек за собой отбор растений на развитие защитных покровов и проводящей системы в наземных условиях.
Цепь событий следовала одна за другой. Развитие непрерывного слоя кутикулы должно было отрицательно сказаться на фотосинтезе, так как препятствовало газообмену растений со средой, поэтому необходимо было обеспечить газообмен и при развитии кутикулы. Это достигается формированием устьица из двух клеток с наружными тонкими и внутренними толстыми стенками, что ведет к изменению просвета щели между ними в зависимости от их тургорного давления — открыванию и закрыванию устьиц. Хотя форма устьиц в ходе эволюции подвергалась изменениям (известны 10 типов устьиц по строению), принципиальный механизм их регуляции остается неизменным у всех наземных растений (включая и механизм поступления воды в клетки устьиц). В ходе эволюции растений на суше менялось несколько и строение самого устьичного аппарата (по мере аридизации климата) в сторону неравномерного увеличения толщины стенок и возникновения побочных клеток. При отсутствии необходимости устьичные клетки редуцируются (риниофиты, растения-паразиты, подводные листья). Из 10 известных типов устьичного аппарата наиболее древним признан анемоцитный, который конвергенггно может появляться и на поздних этапах эволюции (Ал. А. Федоров, 1978).
Некоторые наиболее древним считают пароцитный тип устьиц. Устьичный аппарат отличается полиморфизмом в пределах семейств, рода и даже у отдельного растения. Так, у амарантовых описаны почти все типы устьичного аппарата, характерные для цветковых. Местоположение устьиц на растении (побег, верхняя и нижняя поверхности листовой пластинки) также влияет на структурный тип устьиц. Вероятно, в функциональном отношении все типы устьиц довольно сходны, хотя их разнообразие обусловлено особенностями замыкающих клеток и листовой пластинки. Быть может, в этом также заключается один из парадоксов изменения формы, на что обращают внимание С. В. Мейен, 1988; В. А. Красилов, 1988.
Развитие устьиц в ходе эволюции не привело бы к заметным успехам в борьбе за существование, если бы оно не шло параллельно с усилением проводящей функции растений. Поэтому с повышением роли устьиц в жизни растений происходит постепенное развитие активности проведения воды вверх по стеблю в сторону листьев. Как раз с выходом растений на сушу возникает необходимость уменьшения потерь воды и усиления ее движения. В этом направлении совершенствовался проводящий аппарат: выделялись сосуды и трахеиды, возникала поровость, повышалась роль мертвых клеток в восходящем токе и т. д. Без таких изменений развитие устьиц сыграло бы отрицательную роль в водообмене растений. Сосуды и их членики стали неотъемлемой частью ксилемы только у цветковых, хотя первые из них были бессосудистыми. Ксилема состояла из трахеид лестничного типа. С увеличением сухости воздуха во флоре снижается процент растений с сосудами лестничной перфорации. В результате усовершенствования проводящей системы в нескольких направлениях скорость ксилемного транспорта достигает более 10 м/ч, а флоэмного — 1 м/ч.
Такая скорость ксилемного транспорта не могла быть обеспечена проведением воды и растворенных веществ по живым клеткам, что сопряжено с большими энергетическими затратами. Поэтому в ходе эволюции растений наблюдается постепенное удлинение пути по мертвым клеткам. Как раз с возникновением трахеид, особенно сосудов (наземные растения), с образованием сосудов лестничной перегородки усиливается функция передвижения воды и веществ, обеспечивается возможность переброски их на значительные расстояния.
У первых цветковых описаны лишь трахеиды лестничного типа.
Несмотря на разнообразие сосудов у цветковых, они образуют морфологический ряд перехода от члеников с многочисленными лестничными перфорациями до члеников с простыми перфорациями на поперечных стенках. Специализация сосудов и соответственно усиление их проводящей функции в ходе эволюций покрытосеменных были связаны с уменьшением длины, утолщением стенок, расширением просвета, округлением пор, возникновением поперечной стенки, уменьшением числа перекладин и увеличением ширины сквозных отверстий сосудов. Эти и другие структурные изменения привели к уменьшению сопротивления току жидкости и обеспечили ее подъем на большую высоту.
В течение эволюции совершился переход ситовидных клеток (богатых порами) в членики ситовидных трубок. В результате было достигнуто усиление флоэмного транспорта. Элементы, аналогичные флоэмным, появляются уже у некоторых водорослей. Однако с выделением удлиненных клеток, снабженных ситовидными отверстиями, способность к проведению органических веществ усиливается. Дальнейшая интенсификация флоэмного транспорта оказалась возможной благодаря разрушению ядра у ситовидных клеток. Последнее имело бы отрицательные функциональные последствия, если бы ситовидные клетки были лишены клеток-спутниц, отличающихся интенсивной жизнедеятельностью. Отрицательные последствия потери ядра компенсируются усилением взаимодействия ситовидных клеток и их спутниц.
Различные виды и таксоны растений отличаются по уровню организации проводящей системы.
Как указывает А. Л. Курсанов (1976), различия в проводящей системе разных таксонов не всегда правомерно рассматривать как промежуточные этапы общей дифференциации, скорее они представляют «отдельные ветви эволюционного древа». Автор так подытоживает представления об эволюции системы дальнего транспорта в растительном мире: «Вероятно, исходным было передвижение веществ в недифференцированных клетках с помощью конвекционных токов цитоплазмы, что явилось существенным шагом вперед по сравнению с диффузией и что и теперь еще часто остается основным механизмом внутриклеточного перераспределения веществ. Однако для осуществления транспорта на далекие расстояния этот процесс должен был быть еще более ускорен, что и привело к возникновению экстраподвижности в виде той, которую можно наблюдать в клетке ацетабулярии, в пыльцевой трубке или в корневом волоске».
Сказанное можно подытожить так: у одноклеточных водорослей транспорт веществ ограничен обменом между клеткой и средой, а также между органеллами. У многоклеточных низших растений появляются межклеточные потоки веществ, приводящие к трофической интеграции тканей. У высших сосудистых формируются механизмы быстрого и однонаправленного движения веществ между органами. Такое эволюционное усложнение транспорта веществ связано с возникновением специализированных структур для обеспечения потока веществ — симпласта, апопласта, плазмодесм и переходных клеток (для перекачки веществ из алопласта в симпласт и обратно). В эволюционном ряду от мхов до цветковых растений симпласт усложняется образованием ситовидных трубок, а апопласт упрощается путем образования мертвых сосудов (J. Moorby, 1981).
В связи с этим объяснимы причины различий растений подлине мелких жилок на листьях. Так, при площади листа дыни 1,68 дм2 , их длина составляет примерно 115 м. Изучение около 730 видов растений показало, что переход от деревьев к травам сопровождается изменением плазмодесменных связей между клетками мезофилла и пучка, увеличением числа поглощающих клеток, разным сочетанием симпластного и апопластного путей (Ю. В. Гамалей, 1994). Соответствующие структурные различия способствуют усилению загрузки флоэмы и изменению состава флоэмного экссудата.
Ухудшение водообмена растений в связи с аридизацией климата Земли в поздних геологических эпохах потребовало изменения структуры уже самого организма как целостной системы для восстановления равновесия между поступлением и отдачей воды. Уже у однолетних трав преобладает подземная (всасывающая) поверхность над надземной (испаряющей). Только такой тип организации индивидуума мог обеспечить жизнедеятельность растений в новых условиях даже при наличии совершенной проводящей системы (В. О. Казарян, 1990).
Связь формы и функции в ходе эволюции растений можно продемонстрировать и на других примерах. Заслуживают упоминания молекулярный и клеточный уровни. Наличие сложной внутренней организации клетки и органелл (многочисленных крист, гран, тилакоидов и мембран) обеспечивает непрерывность протекания энергетических и синтетических процессов. Причем и в онтогенезе по мере усложнения внутренней организации органелл их функциональные возможности постепенно усиливаются. На примере химической изомерии биологических молекул (еще со времен Л. Пастера) показана возможность значительного изменения функциональной активности молекул. У молекулы одного и того же фермента может существовать несколько изомеров, отличающихся по некоторым деталям строения («правые» и «левые» формы) и физиологической активности.
Для дальнейшего углубления рассматриваемого вопроса представляют интерес данные об изменении симпластических контактов в онто- и филогенезе растений (Ю. В. Гамалей, 1994; М. В. Пахомова, 1981). Специализация плазмодесм и плотность их распределения возрастают от мезофилла к ситовидным элементам. У бурых водорослей развитие симпластических связей ограничено формированием плазмодесменного поля, у голосеменных и мхов — формированием ситовидных полей, у покрытосеменных — настоящих ситовидных пластинок.
На примере эволюции корня также показано, что усиление его функций шло параллельно нарастанию внутренней дифференциации и гетерогенности клеток верхушки, образованию многоклеточной паренхимы, системы межклетников, поясков Каспари (пересекающих радиальные и поперечные стенки эндодермы) и т. д. (М. Ф. Данилова, 1974). С усилением гетерогенности клеток верхушки корня возрастала избирательность поглощения минеральных элементов.
Особенности транспорта ионов в растениях определяются спецификой структур, что прослеживается в иерархическом ряду от архитектуры биологических мембран до организации ценоза (Ц. Кларксон, 1978). В растениях имеются механизмы, контролирующие внутреннюю ионную среду (меняющие молекулярную организацию мембраны), взаимосвязь между ее элементами и метаболическим аппаратом клетки и структурной организацией самого растения.
Соответствие структуры и функции видно и на примере растений с С3— и С4-путями фотосинтеза. Здесь же отметим, что С4-путь имеет специфические структурные особенности, пространственное и временное разделение мест первичной фиксации CO2 и, его дальнейшее вовлечение в фотосинтез.
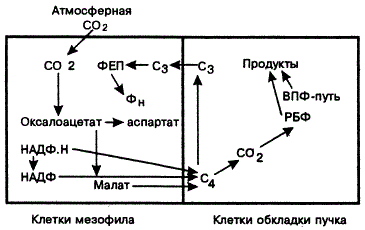
Не только в филогенезе, но и в онтогенезе есть соответствие между структурой хлоропластов и интенсивностью фиксирования в них CO2. Вообще адаптации растений к новому световому режиму в онтогенезе сопровождаются перестройкой листа на тканевом и клеточном уровнях, числа пластид и т. п. (А. Т. Мокроносов, 1981; А. А. Шахов, 1993). Отдельные элементы мезоструктуры фотосинтетического аппарата взаимосвязаны в онтогенезе. Хотя строение ФА тесно коррелирует с его функцией (Дж. Эдвардс, Д. Уокер, 1986), не всякие структурные и функциональные изменения возникают одновременно и оказываются выгодными для их носителя. Более того, возникающие в ходе эволюции и сальтационно крупные макромутации первично не всегда оказывались функционально более значимыми, чем имеющиеся. Однако отбор часто «попустительствовал» их фиксации ввиду нейтральности или слабой адаптивной значимости. В макроэволюции высших растений морфогенез и семифилогения (онто- и филогенез) структур часто предшествовали функциональному совершенствованию индивидуумов и популяций (С. В. Мейен, 1987).
Обращалось внимание и на неполное соответствие плана организации растений принципу адаптивности. Так, К. Уордлоу (С. Wordlaw, 1965) приводит случаи, когда развитие прилистников, зазубренность пластинки и неравномерность ее окраски, утолщенность черешка и сложное строение цветка не всегда отвечают указанному принципу.
Удивительны примеры взаимосвязи формы и функции для биологических молекул. Так, для гиббереллинов показано, что изменение конфигурации и степени насыщения тетрациклической молекулы (метильными и карбоксильными группами) в разных положениях резко меняет их физиологическую активность. Число гиббереллинов, описанных по этому принципу, превышает 60. Некоторые из них характерны только для грибов, другие — для высших растений, третьи — для тех и других. Структура (или организация) должна быть признана характернейшей чертой живого (А. Сент-Дьерди), как и функция. В то же время функциональные особенности имеют некоторые опережающие возможности для изменения. Так, на примере системы биосинтеза хлорофилла продемонстрирована возможность регуляции размеров фотосинтетической единицы в зависимости от условий — состояния функционирования самой молекулы.
Один из примеров влияния функциональных преобразований на структуру органов — укорененные листья. Их черешки после длительного самостоятельного культивирования приобретают сходство по строению со стеблем, в них усиливаются проводящая и механическая функции (Н. И. Дубровицкая).
Структура и функция — неотъемлемые свойства живой природы, они взаимосвязаны в онто- и филогенезе. Любой орган обладает разнообразием функций. Если из множества функций корня (проведение веществ или их запасание, образование придаточных почек, прикрепление, синтез и т. п.) одна получит преимущественное развитие, то строение корня в филогенезе изменится сообразно этой функции. В то же время отбор растений может происходить по количественному выражению одной и той же функции. Такие изменения способствуют изменению и структуры органа.
Соответствие строения органа его функциям не следует понимать узко (в отношении любых морфологических особенностей), оно относится к системе органов и организму в целом. Теория отбора не исходит из признания пользы любых особенностей организации.
Отбор способствует возникновению фенотипов, у которых достигается использование первичных механизмов какого-либо процесса в адаптивных для них структурах. С переходом на другой структурный уровень происходит усиление процесса при сохранении первичных механизмов.
ДИФФЕРЕНЦИАЦИЯ ТЕЛА У РАСТЕНИЙ
↑
СПЕЦИАЛИЗАЦИЯ СТРУКТУР (ОРГАН)
↑
ВЗАИМОДЕЙСТВИЕ КЛЕТОК (ТКАНЬ)
↑
ЭУКАРИОТИЧЕСКАЯ КЛЕТКА
↑
ПЕРВИЧНЫЕ МЕХАНИЗМЫ (ПРОКАРИОТЫ)
В итоге наблюдается заметная функциональная дивергенция растений, что также зависит от таксономического и экологического рангов вида. Значение подобной дивергенции для эволюции растений исключительно.
Проблема взаимосвязи структуры и функции должна быть разработана применительно к разным уровням живой природы. Так, в основе фотосинтеза лежат общие физико-химические механизмы и реакции, протекающие на мембранах. Однако в ходе эволюции фотосинтез менялся на уровне организма, популяции и растительности в целом. Отсюда и изменения структуры, и функции фотосинтетического аппарата на разных уровнях его организации возникали в разное время и с помощью различных механизмов.
В рамках конкретного физиологического проявления необходимо рассматривать его эволюцию на всех уровнях организации. Однако физиологи растений традиционно взаимосвязь структуры и функции ограничивают организменным и клеточным уровнями, так как организм представляет основной предмет их интереса. В то же время очевидно, что этим не может быть исчерпан сложный вопрос о возникновении того или иного физиологического явления. Следовательно, для понимания глобального значения «растительной жизни» необходимо исследовать ее основные параметры с точки зрения их реализации на всех уровнях организации жизни.
Фотосинтез возникает в результате эволюции надорганизменных уровней для выполнения растениями космической функции. Из этих соображений исходят при оценке роли тех или иных организмов на Земле. Функция фотосинтеза на уровне надорганизменных образований видна при оценке общей биомассы, ежегодной и удельной продуктивности фитопланктона и фитобентоса в Мировом океане. Половина от ежегодной продукции кислорода растениями (3,2 ∙ 1011 т) приходится на Мировой океан. Для создания современного фонда кислорода биосферы при этом водорослям понадобилось бы 3700 лет (А. И. Перельман, 1979). Что прежде всего определяет эволюционное назначение фотосинтеза в поддержании жизни на планете.
Только с этих позиций проясняются некоторые факты изменения структуры и функции организма и в онтогенезе. Так, мы знаем о падении обменных (основных) процессов растений по мере старения организма и органа, т. е. с увеличением общей массы тела, всех его структурных компонентов.
На примере популяций Cyctoseira crinita показано, что молодые особи в мелководьях и условиях высокой подвижности воды обеспечивают поток 1800 г углерода/м2 дна, тогда как взрослые особи, обитающие на большой глубине и при спокойной воде, — 200—300 г углерода/м2 дна (К. М. Хайлов, З. П. Парчевский, 1983). Причины таких изменений могут быть различными, в том числе и как проявление в онтогенезе филогенетических тенденций развития.
Влияние структуры на функцию не всегда однозначно. Так, большое количество ДНК в ядре значимо для продолжительности митоза, мейоза и жизненного цикла. Однако было бы неверно связывать содержание ДНК в ядре только с указанными событиями.