Любая биологическая система, будь то клетка или организм, биологическая семья (наподобие пчелиной семьи) или целая генетическая популяция, обладает системными адаптивными механизмами, с помощью которых она поддерживает свое существование.
Известно, что организму свойствен целый ряд приспособительных механизмов, позволяющих ему сохранять свою внутреннюю среду и противостоять резким колебаниям внешней среды (физиологический гомеостаз). Физиологический гомеостаз, по-видимому, может действовать и на клеточном уровне посредством клеточных адаптационных физиологических механизмов (клеточный гомеостаз).
В панмиктической популяции имеются адаптационные механизмы, обеспечивающие ее существование. В основе существования панмиктической популяции как единой системы лежат в первую очередь генетические механизмы, осуществляющиеся благодаря свободному скрещиванию разнополых особей и постоянному обмену наследственной информацией между особями популяции.
Процессы, обеспечивающие способность популяции сохранять свою генетическую структуру в ответ на воздействие факторов внешней среды, недавно было предложено М. Лернером называть генетическим гомеостазом. Та же идея была сформулирована С. С. Четвериковым еще в 1926 г.: «Видовое сообщество в условиях свободного скрещивания представляет собой устойчивый агрегат, внутри которого в самих условиях свободного скрещивания заложен аппарат стабилизации численных отношений составляющих его аллеломорфных пар (законы Гарди и Пирсона)».
В основе существования генетического гомеостаза на уровне популяции лежат механизмы, обеспечивающие ее способность приспособительно поддерживать свой генетический состав. К этим механизмам относятся:
- поддержание равновесного состояния популяции по генотипическим частотам в соответствии с формулой Гарди — Вайнберга,
- поддержание гетерозиготности и полиморфизма,
- поддержание определенного темпа и направления мутационного процесса.
Механизм поддержания равновесия в популяции по генотипическим частотам был рассмотрен выше, здесь же остановимся на механизме обеспечения гетерозиготности и полиморфизма.
Мы говорили о том, что под внешним относительным однообразием панмиктической популяции скрывается огромное генетическое разнообразие.
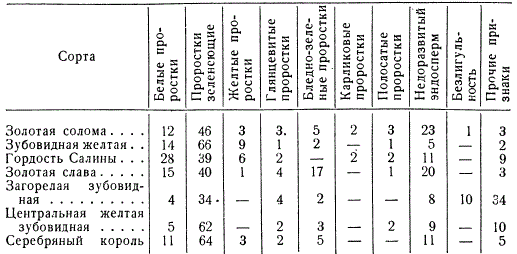
Количество растений, гетерозиготных по различным рецессивным мутациям, установленным для некоторых сортов кукурузы (в %)
Показан высокий процент гетерозиготных растений по различным мутациям у различных сортов кукурузы. При исследовании популяции ржи, состоящей из 167 растений, у 6% растений были обнаружены хромосомные перестройки в гетерозиготном состоянии. При изучении популяций львиного зева было установлено, что до 15% растений являются носителями различных мутаций. Исследуя природные популяции дрозофилы, С. С. Четвериков показал высокую их насыщенность различными мутациями в гетерозиготном состоянии.
Популяции содержат огромное количество разнообразных рецессивных мутаций, различных хромосомных перестроек, концентрации которых меняются в зависимости от размера популяций, условий внешней среды и темпа мутационного процесса.
Насыщенность популяций мутациями обеспечивает резерв наследственной изменчивости. При перемене условий внешней среды и изменении направления отбора резерв мутаций в гетерозиготном состоянии позволяет популяции в более короткие сроки приспособиться к изменившимся условиям за счет изменения генетической структуры. Следовательно, гетерозиготное состояние особей популяции обеспечивает ее приспособительную пластичность. Кроме того, гетерозиготы, как правило, имеют более высокую жизнеспособность, чем гомозиготы. У них шире норма реакции генотипа, т. е больший диапазон приспособительных возможностей, чем у гомозигот, что и обеспечивает им селективное преимущество.
Ч. Дарвин первым обнаружил явление биологической полезности скрещивания. В последующем было установлено, что гетерозиготное состояние генов нередко ведет к повышению мощности и жизнеспособности гибридных организмов по сравнению с родительскими формами. Это явление было названо гибридной силой, или гетерозисом. Напротив, близкородственное скрещивание (инбридинг), повышающее гомозиготность особей в поколениях, часто вызывает депрессию в потомстве, или вырождение. Таким образом, гетерозиготность в популяции является одним из важных механизмов генетического гомеостаза.
Другим адаптационным генетическим механизмом, направленным на обеспечение целостности популяции как единой системы, является наличие в ней наследственного полиморфизма. Полиморфизмом популяции называют существование в ней целого ряда форм, определяемых генотипической изменчивостью и воспроизводящихся при размножении.
Если генотипические различия сопровождаются фенотипическими различиями и гетерозиготы имеют адаптивное преимущество, то в популяции при отборе в пользу гетерозигот создается сбалансированный полиморфизм. Сбалансированным полиморфизмом и называется воспроизведение в популяции из поколения в поколение классов особей, различающихся генотипически и фенотипически.
Наличие сбалансированного полиморфизма не означает, однако, жесткой фиксации генотипических частот. Н. В. Тимофеев-Ресовский и Я. Я. Лус на протяжении многих лет проводили исследование сходных полиморфных популяций божьих коровок (Adalia bipunctata), включавших особей двух основных классов — с красной и черной окраской надкрыльев, и из года в год в данных популяциях наблюдалась одна и та же картина: осенью преобладали черные жуки, тогда как после перезимовки резко возрастала частота красных жуков.
Это, казалось бы, простое наблюдение позволило вместе с тем прийти к весьма существенным заключениям. Во-первых, приспособительная ценность каждой из особей не является постоянной и меняется при изменении условий (в зависимости от сезона); во — вторых, наличие полиморфизма в популяции обеспечивает возможность регулировки ее состава за счет приспособительной динамики соотношения частот различных классов особей (например, Аа и аа); и, в-третьих, сохранение в течение многих лет полиморфного состава популяции и предотвращение полной элиминации какого-либо из классов может указывать на наличие механизма отбора в пользу гетерозигот. Выводы, полученные в данном исследовании, нашли затем подтверждение при анализе природных популяций улиток, бабочек, богомолов, хомячков и других объектов. Приведем пример с рыбами.
У морских лососевых рыб, нерестящихся в северных реках, существуют две формы самцов. Одни самцы после ската растут в море и приобретают крупные размеры тела, достигая 100 см в длину. К местам нереста они приходят в возрасте 5—7 лет вместе с самками. Другие самцы — карликовые достигают половозрелости в возрасте 2—3 лет; длина их не более 15 см. Причем эти самцы живут только в реке. Наряду с морскими собратьями они принимают участие в оплодотворении икры и воспроизводят как нормальное потомство, которое затем претерпевает скат в море и возвращается на нерест после интенсивного роста, так и карликовых самцов, остающихся в реке.
Как предполагала Н. В. Европейцева, присутствие карликовых самцов характерно для тех речных популяций, в которых нарушено соотношение полов при возврате рыб на нерест. Таким образом, их существование более надежно обеспечивает необходимое соотношение полов для воспроизводства популяции.
Классическим примером полиморфизма служит разделение функций между различными формами у общественных насекомых: пчел, муравьев, термитов. Причем появление таких форм связано у них с особенностями полового процесса и мейоза и регулируется также онтогенетическими механизмами.
У растений еще Ч. Дарвином было изучено явление гетеростилии, которое также представляет пример полиморфизма. Первоцвет (Primula vulgaris) встречается в двух формах, различающихся по строению цветков. Примерно у половины растений рыльце пестика в цветке выступает наружу, а пыльники сидят на коротких тычиночных нитях и спрятаны в трубке венчика. У другой половины растений наружу выступают пыльники, а рыльце спрятано в трубке венчика. Такое строение цветков является надежным приспособлением для обеспечения перекрестного опыления насекомыми. При принудительном самоопылении растений, имеющих цветки с длинным столбиком и короткими тычинками, в потомстве возникают все растения такого же типа. При самоопылении растений, имеющих цветки с длинными тычинками и коротким столбиком, в первом же поколении наблюдается расщепление в соотношении 3 : 1 (3 с длинными тычинками и коротким пестиком и 1 с короткими тычинками и длинным пестиком). При перекрестном опылении осуществляется расщепление в соотношении 1:1. Отсюда ясно, что в основе наследственного определения механизма гетеростилии лежит расщепление по паре аллелей одного гена S. В природе все растения с длинными тычинками и коротким пестиком постоянно гетерозиготны (Ss), поскольку переопыление гомозиготных растений одного типа между собой не может осуществляться. В результате все время происходят возвратные скрещивания на рецессивную форму (ss), и, таким образом, поддерживается численно равное соотношение обеих форм.
Рассмотрение явления полиморфизма убеждает в том, что наличие его в популяции оказывается необходимым для существования последней. Естественный отбор закрепляет существование полиморфизма, контролируя численное соотношение необходимых форм в каждом поколении. Такой вид полиморфизма называют в литературе сбалансированным полиморфизмом.
Полиморфизм является механизмом поддержания генетической популяции как единой системы. Поэтому его можно рассматривать как проявление генетического гомеостаза, развившегося в процессе эволюции, в результате действия естественного отбора.