Еще в 1911 г. Р. Гольдшмидт и ряд других зоологов показали, что при реципрокных скрещиваниях различных географических рас некоторых насекомых в потомстве наряду с самцами и самками возникают формы, имеющие постепенные переходы в отношении первичных и вторичных половых признаков от мужского пола к женскому, т. е. формы, которые теперь называют интерсексами.
Организмы являются генетически бисексуальными в том смысле, что в их генотипе имеются гены обоих полов, но соотношение этих генов, а возможно, и характер их действия у мужских и женских особей различны. Теория Р. Гольдшмидта строилась исходя именно из таких представлений.
Физиологическая теория определения пола оснозывалась на исследовании интерсексов, получаемых путем реципрокного скрещивания различных географических рас непарного шелкопряда (Limantria dispar). При этих скрещиваниях не происходит изменения в соотношении половых хромосом и наборов аутосом — интерсексы являются диплоидными по всем хромосомам. Поэтому возникающие в потомстве промежуточные в половом отношении формы представляют собой диплоидных интерсексов.
Для скрещивания Гольдшмидт избрал две географические расы непарного шелкопряда — европейскую и японскую, различающиеся по половой потенции: европейская раса была отнесена по этому признаку к «слабой», а японская — к «сильной». Оказалось, что в том случае, когда самки европейской расы скрещиваются с самцами японской, в F1 развивается интерсексуальность женского типа.
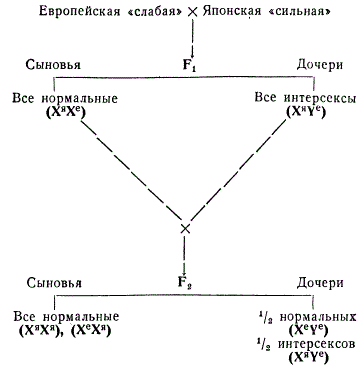
У бабочек непарного шелкопряда гетерогаметным полом (XY) является женский, а гомогаметным (XX) — мужской. Можно предоставить, что интерсексуальность и в этом случае объясняется балансом двух тенденций: женской, определяемой Y-хромосомой, и мужской, определяемой Х-хромосомой. При этом играет роль «сила» и «слабость» этих факторов у разных рас. Х-хромосомы непарного шелкопряда в отличие от дрозофилы содержат гены, определяющие мужской пол, а гены женского пола локализуются предположительно в Y-хромосоме. У европейской расы «женская» тенденция Y-хромосомы «сильнее» мужской тенденции Х-хромосомы, вследствие чего при размножении внутри расы особь XY постоянно развивается в самку, а XX — в самца. То же самое наблюдается и у японской расы, дающей внутри себя нормальное соотношение полов без интерсексов.
Но X- и Y-хромосомы разных рас обладают разной силой в отношении определения пола. В данном примере Xя (Xя — хромосома японской расы) имеет более сильный мужской фактор, так же как и Yя — женский (Yя — хромосома той же расы), по сравнению с Xе— и Yе-хромосомами европейской расы. Поэтому у дочерей F1 рассматриваемого скрещивания мужской фактор Х-хромосомы японской расы (Xя) способен частично подавить женский фактор Y-хромосомы европейской расы (Ye) и вызвать интерсексуальность, втором поколении половина дочерей оказывается интерсексами, как они имеют то же соотношение факторов (Xя Ye).
Из рассмотренного опыта следует, что эта гипотеза определения пола также может быть названа гипотезой генного баланса. Результаты проведенных исследований показывают, что организмы действительно бисексуальны — обладают обеими генетическими потенциями в отношении развития пола.
Генетически определяемая бисексуальность обнаружена и у других животных, и у растений. Как мы видели, пол у дремы также определяется соотношением X- и Y-хромосом, но даже женские растения, лишенные Y-хромосомы, имеют мужские потенции. Так, при поражении женских растений головней в цветках развиваются тычинки, т. е. цветки становятся морфологически гермафродитными.