Развитие признаков пола, как и любых других признаков организма, определяется генотипом и факторами внешней среды.
Мы уже показали, что организмы генетически бисексуальны. Именно поэтому процесс дифференциации пола оказывается сложным. Бисексуальная основа организма в принципе позволяет изменять направление его развития, т. е. переопределять пол в онтогенезе.
Теперь в руках биологов имеются уже различные методы — генетические, эмбриологические и цитологические, с помощью которых можно изучать дифференциацию пола и воздействовать на этот процесс с целью переопределения его в индивидуальном развитии животных и растений. Рассмотрим кратко две стороны этой проблемы: дифференциацию пола и переопределение пола в онтогенезе.
Дифференциация пола
Вслед за определением пола следует дифференциация, т. е. развитие половых различий.
Главным критерием пола является формирование воспроизводительной, половой системы и физиологического (биохимического) механизма, обеспечивающего скрещивание.
Зачаточные гонады у эмбрионов животных оказываются двойственной природы или, точнее, индифферентными в половом отношении. Они состоят из внешнего слоя ткани, называемого кортексом (cortex), из которого развивается женская ткань, и из внутреннего слоя, называемого медуллой (medulla), из которого развивается мужская ткань.
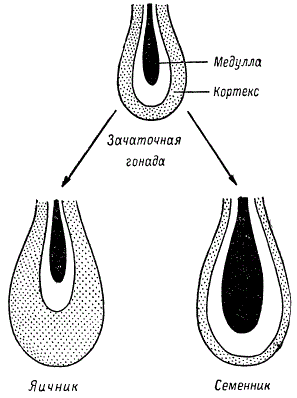
Схема дифференциации гонад в онтогенезе
В ходе дифференциации пола идет развитие одного из зачатков и подавление другого. У мужского пола быстрее развивается медуллярная ткань, которая подавляет деятельность кортикального слоя, в результате гонады превращаются в семенники. У женского пола ускоряется развитие кортикального слоя, в силу чего подавляется формирование медуллярного слоя, и гонады превращаются в яичники. Соответственно этим преобразованиям половых зачатков дифференцируются и половые пути. Вначале закладываются как Вольфовы, так и Мюллеровы протоки, но впоследствии протоки противоположного пола дегенерируют или остаются в зачаточном состоянии.
Половая дифференциация эмбриона человека начинается с образования гонад. У шестинедельного эмбриона, который имеет длину 12 мм, пол еще не различается, гонады внешне одинаковы. У зародыша в 13 мм появляются первые признаки мужских гонад — семенники. Гонады женского эмбриона в это время индифферентны; на неделю позже, чем у мужского эмбриона, в них начинается процесс дифференциации яичников. В 12-недельном возрасте пол эмбриона человека хорошо различается.
Сам процесс дифференциации пола у многих животных обусловлен гормонами, которые выделяются не только эндокринными железами, но и кортикальным и медуллярным слоями полового зачатка, а в последующем — и половыми железами. Эти вещества отличаются по времени их выработки и по характеру своего действия. У особей мужского пола преобладает «мужское вещество» — медулларин, у женского — кортикальный гормон.
Дальнейшая дифференциация пола, особенно развитие вторичных половых признаков, также идет под влиянием различных гормонов. Если у самца млекопитающего удалить семенники, т. е. кастрировать его до наступления половой зрелости, то взрослое животное приобретает некоторые признаки женского пола — оно становится феминизированным. Классическими опытами М. М. Завадовского еще в 20-х годах было показано, что кастрация самок птиц даже во взрослом состоянии приводит к развитию признаков противоположного пола. Кастрированные курочки превращаются по внешнему виду в петухов. Этот процесс может быть обратимым.
Главным действующим началом в дифференциации пола являются гены, контролирующие уровень гормональной секреции мужского или женского направления. Преобладание генов, определяющих мужской пол, в общем балансе приводит к повышению активности мужских гормонов и к дифференциации мужского пола, обратное соотношение генов — к развитию женского пола. Преобладание в онтогенезе гормональной секреции то одного, то другого пола приводит к развитию интерсексуальных форм. По представлению Р. Гольдшмидта, у интерсексов имеются критические моменты гормональной активности, которые определяют «поворот» в развитии пола данной особи. Половые гормоны животных принято делить на три группы: андрогены, эстрогены и гормон желтого тела — прогестерон. Эстрогены и прогестерон являются женскими половыми гормонами, а андрогены — мужскими. Результаты исследований показывают, что гормоны у позвоночных оказывают влияние не только на развитие вторичных половых признаков, но и на гонады. Андрогенные вещества вызывают маскулинизацию яичников, т. е. появление в них мужских половых клеток, а эстрогены — феминизацию семенников, т. е. появление в них женских половых клеток.
На дифференциацию пола у высших растений значительное влияние оказывает действие растительных гормонов — ауксинов.
Переопределение пола в онтогенезе
Наилучшим доказательством наследственной бисексуальности организмов является изменение пола в онтогенезе в естественных или искусственных условиях.
Изучение действия гормонов на дифференциацию пола и переопределение его в онтогенезе является очень важной проблемой генетики, эмбриологии и эндокринологии.
Раньше мы уже говорили о том, что отдельные мутантные гены, например у кукурузы, могут как бы переопределить пол, точнее — превратить однодомное растение в двудомное. У дрозофилы один из аутосомных генов может сдвинуть развитие пола в противоположную сторону. Теперь же мы рассмотрим другую сторону вопроса — ненаследственное, онтогенетическое, изменение, или переопределение, пола. В этом отношении генетики и эмбриологи располагают большим количеством фактов, данных самой природой и полученных экспериментально.
У человека и млекопитающих, как правило, при развитии разнополых близнецов гормональное переопределение пола в эмбриогенезе затруднено тем, что дифференциация пола наступает раньше, чем начинается продуцирование соответствующих гормонов. Однако имеются случаи, когда изменение пола происходит в эмбриогенезе. У крупного рогатого скота иногда рождаются двойни однополые (два бычка или две телочки) и разнополые (бычок и телочка). В случае однополых близнецов они рождаются нормальными. Когда двойни являются разнополыми, то бычки развиваются нормально, а телочки часто оказываются интерсексами: наружные гениталии женского типа, в внутренние органы — мужского. Такие животные были названы фримартинами; они всегда бесплодны. Подобные изменения вызываются тем, что семенники мужского эмбриона иногда раньше начинают выделять мужские гормоны в кровь, которые и вызывают изменение женского эмбриона.
Обычно для иллюстрации переопределения пола в природных условиях приводят несколько классических примеров, например определение пола у морского червя Bonellia viridis. Свободно плавающие личинки этого червя развиваются в самок. Если личинка прикрепляется к хоботку самки или поселяется вблизи нее, то она развивается в самца. Личинка, начавшая дифференцироваться в самца, будучи отделена от самки, изменяет направление дифференциации пола в женскую сторону, и из нее развивается интерсекс. В хоботке самки имеются химические вещества, способные переопределить пол личинок. Предполагается, что такое определение пола является биологически выгодным для данного вида. В генетическом отношении пол здесь также детерминирован, но с равной начальной половой потенцией. Решающим моментом в определении пола оказывается контакт с женской особью.
В роде моллюсков Crepidula имеются виды, особи которых нормально проходят ряд сменяющихся фаз развития: бесполая фаза, мужская, переходная, затем женская. Особи, находящиеся в мужской фазе, после спаривания с особями в женской фазе либо приобретают сидячий образ жизни, либо свободно плавают и в этом случае быстрее, чем прикрепленные формы, переходят к женской фазе.
В последние годы все более привлекает внимание исследователей экспериментальное переопределение пола. Путем воздействия различными гормональными препаратами у ряда животных удается получить полное превращение пола вплоть до способности формирования измененными особями половых клеток противоположного пола. Такие результаты получены у тритона (Pleurodeles waltlii), некоторых лягушек (Xenopus laevis), рыб (Oryzias latipes), птиц и ряда других животных. Кратко остановимся на двух примерах.
Очевидно, что переопределение мужского пола в женский у кур может представить некоторый практический интерес. В 1956 г. Появилось сообщение о том, что обработка эстрогенами куриных яиц в инкубации вызывает полное превращение мужского пола в женский. При этом превращенные в курочек петушки сохраняют признаки самки в последующем развитии. Позднее за границей и у нас в стране была проведена проверка возможности такого переопределения с генетической маркировкой половых хромосом. Опыты В. Б. Савватеева показали, что обработка женским половым гормоном (диэтилстилбестролом) оплодотворенных яиц до инкубации Действительно превращает мужской пол в женский, но только на эмбриональной стадии. При дальнейшем развитии генотип брал верх, и у таких цыплят наблюдалась полная реверсия к мужскому полу.
Один из замечательных примеров полного переопределения пола в онтогенезе получен на аквариумных рыбках в исследовании Т. Ямамото в 1953 г. У многих аквариумных рыбок гетерогаметным полом (X Y) является мужской. В опыт были взяты белые и красные медаки (Oryzias latipes), у которых доминантный ген красной окраски R находится в Y-хромосоме, а его рецессивная аллель r — в Х-хромосоме. Следовательно, белые самки имеют генотип ХrХr, а красные самцы — XrYR. В этом случае самцы всегда будут красными, поскольку в Y-хромосоме находится доминантный ген R.
При указанном типе наследования сыновья всегда будут нести признак отца, если не произойдет кроссинговера между гомологичными участками X- и Y-хромосомы, что иногда имеет место. Предварительно автор проверил наследование этого признака в нескольких поколениях. Скрещивание ХrХr x XrYR неизменно давало белых самок и красных самцов.
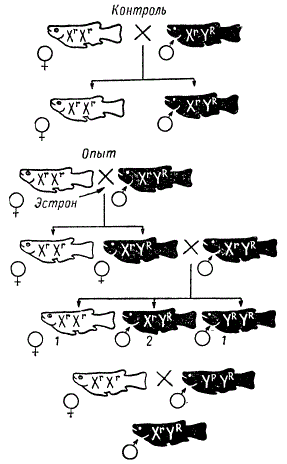
Фенотипическое переопределение пола у рыб
Выклюнувшиеся мальки, пока у них еще не дифференцировался половой зачаток, были разбиты на две группы, которые содержались до 8 месяцев на двух различных диетах:
- нормальное кормление,
- с добавкой женского полового гормона (эстрона или стилбестрола).
В результате оказалось, что все красные рыбки во второй группе, генотипически определяемые как самцы XrYR, по фенотипу оказались самками с нормальными яичниками и с женскими вторичными половыми признаками. Они были способны скрещиваться с нормальными красными самцами.
Скрещивание таких самок с нормальными самцами XrYR x XrYR давало расщепление по полу не 1 : 1, а 1♀(XrXr) : 3♂(XrYR и YRYR).
Этот пример ясно демонстрирует:
- генетическую бисексуальность организмов,
- возможность изменить дифференцировку пола в онтогенезе,
- один из путей искусственной регуляции соотношения полов.