Деятельность камбия у различных растений неодинакова. У однодольных и у некоторых травянистых двудольных камбий функционирует очень короткое время и практически не играет роли в утолщении стебля.
Сосудисто-волокнистые пучки, структура которых в основной своей массе определяется элементами первичных флоэмы и ксилемы, где камбий действует весьма короткое время или совсем не действует, называются закрытыми. Типичные закрытые сосудисто-волокнистые пучки свойственны всем однодольным растениям.
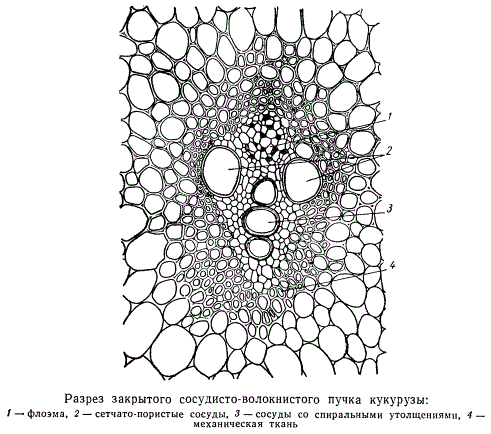
У многих однодольных, например, у злаков, стебли почти совсем не утолщаются, а если несколько и утолщаются, то не за счет увеличения массивности сосудисто-волокнистых пучков, а вследствие разрастания паренхимы стебля. Среди однодольных есть растения, довольно значительно утолщающие стебель по отношению к первоначальной толщине его (драцены, юкки), типичный камбий в этом утолщении также не участвует. Утолщение стебля в этих случаях обусловлено периодическим возникновением в паренхиме меристематических поясов, ближайших к периферии органа. В этих кольцевых поясах возникают новые сосудисто-волокнистые пучки, тоже закрытые.
В противоположность закрытым сосудисто-волокнистым пучкам пучки, имеющие камбий, действующий более или менее продолжительное время, называют открытыми. Обязательным результатом деятельности камбия является образование вторичных флоэмы и ксилемы и вследствие этого утолщение органа. Продолжительность деятельности камбия у различных растений неодинакова. Особенно длительна деятельность камбия у древесных двудольных и хвойных растений, где она продолжается до конца существования самого растения с перерывами в холодные и засушливые периоды. Однако у травянистых двудольных деятельность камбия нередко продолжается короткое время, прекращаясь перед цветением.
Сосудисто-волокнистые пучки травянистых двудольных со слабо проявляющейся деятельностью камбия являются тоже по существу закрытыми. Весьма незначительно утолщаются сосудисто-волокнистые пучки в листьях, и только в жилках первых порядков деятельность камбия иногда бывает заметна. Сосудисто-волокнистый пучок, образованный таким образом, что наружную его
часть занимает флоэма, а с внутренней стороны примыкает ксилема, называется коллатеральным. Есть пучки и другого типа. У некоторых растений (тыквенные, пасленовые) в пучках ковнутри от ксилемы расположена еще внутренняя флоэма. Сосудисто-волокнистые пучки с двумя участками флоэмы, наружным и внутренним, называются биколлатеральными. Если флоэма в сосудисто-волокнистом пучке расположена так, что ее окружает ксилема, а флоэма находится внутри (иногда наоборот), пучки называются концентрическими. В корнях однодольных, а также в молодых корнях двудольных растений пучки ксилемы чередуются с пучками флоэмы, образуя круг и располагаясь по радиусам этого круга. Такое расположение проводящих тканей называется радиальным.
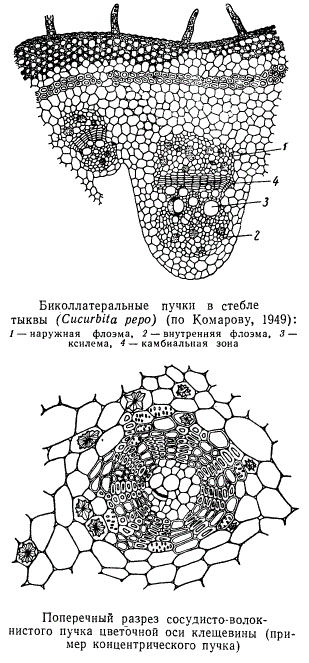
Сосудисто-волокнистые пучки в каждом органе растения нередко образуют довольно сложную сеть, построенную вполне закономерно. Особенно густая сеть пучков бывает в некоторых плодах. Всем известна люффа из семейства тыквенных, употребляемая как мочалка, — не что иное, как сеть сосудисто-волокнистых лучков плода, освобожденного от мякоти.
Отложенные камбием клетки вторичных флоэмы и ксилемы сначала расположены строгими радиальными рядами. Затем в зависимости от того, разнообразны или нет дифференцирующиеся вторичные элементы, строгий радиальный порядок расположения сохраняется или нарушается. При однообразии анатомических элементов ксилемы и флоэмы они будут располагаться рядами, идущими от сердцевины до коры. В эти ряды входит и первичная ксилема.
Клетки первичной флоэмы и особенно ситовидные трубки нередко облитерируются, их полости сжимаются.
Группа анатомических элементов, состоящая из радиально расположенных первичной ксилемы, вторичной ксилемы, узкой полоски камбия и вторичной флоэмы с остатками первичной флоэмы, называется элементарным пучком.

Если же вторичные элементы сосудисто-волокнистых пучков разнообразны, то радиальное расположение частей элементарных пучков смещается. Особенно значительные нарушения в порядок расположения элементов пучка вносят сосуды, которые бывают весьма разнообразных размеров.
Наряду с увеличением массивности сосудисто-волокнистых пучков в радиальной плоскости происходит их увеличение и в тангентальном направлении. Из меристемы, занимающей промежутки между первично возникшими сосудистыми пучками, и меристемы, образующей так называемые первичные сердцевинные лучи, постепенно дифференцируется межпучковый камбий, соединяющий отдельные полоски пучкового камбия в сплошное камбиальное кольцо.
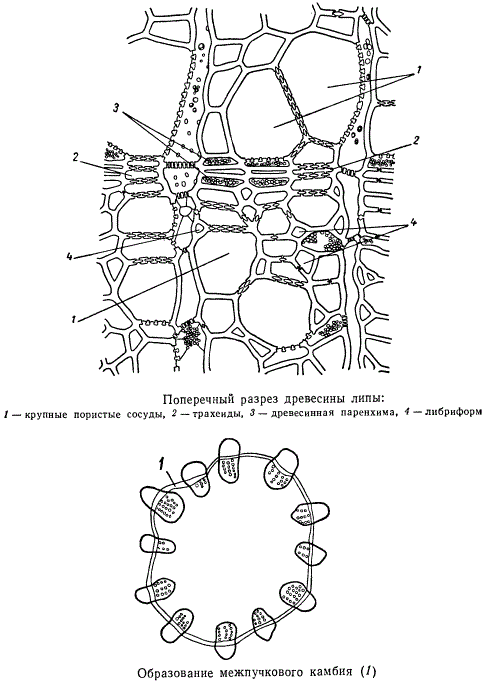
С появлением кольца вторичной меристемы массив основной паренхимы органа отчетливо разделяется на две зоны: периферическую и центральную. Периферическая называется корой (первичной), центральная — сердцевиной, причем между корой и сердцевиной расположено кольцо из сосудисто-волокнистых пучков. У многих растений, особенно у древесных, первичная кора полностью сбрасывается при возникновении перидермы, остается лишь вторичная кора. Собственно, настоящая первичная кора состоит лишь из одной периферически расположенной паренхимы, вторичная же кора обязательно содержит в своем составе элементы лубяной части сосудисто-волокнистой системы.
С возникновением межпучкового камбия, соединяющего сосудисто-волокнистые пучки, все кольцо пучков начинает постепенно менять свой вид. Прежде всего существующие пучки заметно расширяются в тангентальной плоскости. Это расширение обусловлено тем, что в пределах каждого пучка образовавшийся камбиальный слой разрастается за счет радиальных делений.
Увеличившийся ряд камбиальных клеток, разумеется, откладывает более широкую полосу вторичных элементов сосудисто-волокнистого пучка. Если сравнить рисунки, можно отчетливо представить себе расширение сосудисто-волокнистого пучка в тангентальной плоскости массива. На обоих рисунках изображен один и тот же пучок, но на различных уровнях вдоль по стеблю. На рисунке разрез пучка сделан на более высоком уровне стебля по сравнению с пучком, изображенным на другом рисунке. Чем ниже вдоль по стеблю расположен участок сосудисто-волокнистого пучка, тем отчетливее выражены результаты деятельности камбия, тем дальше продвинулся процесс формирования пучка.

На указанных рисунках изображены поперечные разрезы сосудисто-волокнистого пучка рами. Обратим внимание только на вторичную ксилему, в данном примере наиболее четко выраженную. Вторичная ксилема пучков стебля рами в начале своего возникновения содержит весьма мало сосудов, тем более широкопросветных, и состоит преимущественно из либриформа и трахеидоподобных клеток. Вследствие этого вторичная ксилема стебля рами вначале очень однородна, а потом в ней появляются широкоиросветные сосуды. В более верхнем участке сосудисто-волокнистого пучка, в радиальной плоскости, отложилось 9 рядов клеток вторичной ксилемы, а на границе с камбиальной зоной, т. е. в тангентальной плоскости, — 25 рядов. В более нижнем участке пучка, в радиальной плоскости, находится 16 рядов клеток вторичной ксилемы, а на границе с камбием, в тангентальной плоскости, — 27 рядов. Камбий, отложив в нижнем участке пучка 7 лишних слоев ксилемы, увеличил также число радиальных рядов элементов ксилемы в 2 раза. Кроме того, клетки ксилемы значительно разрослись в своих поперечниках. Таким образом, параллельно с ростом пучка в радиальном направлении происходит увеличение его и по периферии.
Обратимся опять к перемычкам вторичной меристемы между первично возникшими пучками — межпучковому камбию.

Межпучковый камбий является тем слоем, где возникают новые сосудисто-волокнистые пучки. Сначала там, где должен возникнуть пучок, намечается группа мелких клеток с зернистым содержимым, которые возникают путем более усиленного местного деления соответствующих клеток перемычек и имеют сходство с клетками прокамбия. Затем из этого участка меристемы дифференцируются элементы сосудисто-волокнистого пучка с прослойкой камбия между ними.
Вновь возникший пучок развивается как в радиальном, так и в тангентальном направлениях описанным выше способом. Следовательно, образовательная ткань, подобная прокамбию, может возникать не только из первичной меристемы. Вообще такая меристема образуется даже из очень старых паренхимных клеток, если последние еще не потеряли своей потенциальной эмбриональности и если создаются соответствующие условия (например, при ранениях растительных органов, подрезках, прививках и т. п.).
Вполне развившийся сосудисто-волокнистый пучок нельзя представлять себе как нечто раз и навсегда образовавшееся и структурно неизменное. В процессе развития всякого сосудисто-волокнистого пучка происходит не только новообразование анатомических элементов, но и их отмирание и удаление из системы элементов, слагающих пучок.
Ксилема однодольных растений и первичная ксилема двудольных, образующиеся без участия камбия, весьма однородны по своему составу. Разнообразие анатомических элементов ксилемы возникает там, где они образуются за счет камбиальной деятельности. Наиболее отчетливо разнообразие элементов вторичной ксилемы выступает в тех случаях, если в непосредственном соседстве одновременно встречаются сосуды и механические элементы (либриформ, заменяющие волокна, волокнистые трахеиды и т. п.) или элементы древесинной паренхимы. Такую структуру вторичной ксилемы имеют стебли рами. Широкополостные сосуды обычно резко выделяются в комплексе анатомических элементов, составляющих вторичную ксилему. Анатомические элементы проводящей системы различных органов растения и особенно стебля могут быть весьма разнообразными не только по своей морфологии и калибру, но и по длине. При этом значительные колебания в длине проявляются как среди разнородных анатомических элементов, так и среди элементов одного и того же типа. Последнее обстоятельство особенно важно для представления сущности структуры проводящей системы и соотношения составляющих эту систему элементов.
Количественно-анатомические соотношения однородных элементов вторичной ксилемы подробно были изучены у различных древесных растений.
Длина клеток вторичной ксилемы обусловливается, во-первых, длиной камбиальных клеток, и во-вторых, разрастанием производных камбиальных клеток, осуществляющимся в процессе дифференциации их в постоянные анатомические элементы. Основные вариации длины, наблюдаемые, например, среди трахеид вторичной ксилемы, прямо связаны с подобными же вариациями в длине камбиальных клеток. Клетки камбия по своей длине весьма разнообразны в различных частях растения или же отдельного его органа. Кроме того, у различных групп сосудистых растений, притом в зависимости от условий их местообитания, длина камбиальных клеток также разнообразна. В раннем возрасте у хвойных и древесных двудольных в развитии стебля наступает период, в течение которого длина элементов древесины увеличивается довольно быстро. Продолжительность этого периода, а также быстрота удлинения анатомических элементов древесины за это время более или менее значительно варьирует не только у различных групп растений, но и у представителей одного и того же вида, а также на разной высоте стебля одного и того же растения. Различные типы анатомических элементов древесины, трахеиды, волокна либриформа и отдельные членики сосудов ведут себя по-разному. Трахеиды в зрелой древесине голосеменных в среднем более чем в два раза длиннее всех разнообразных анатомических элементов вторичной ксилемы двудольных.
Длина прозенхимных анатомических элементов древесины в течение некоторого периода от начала развития растения увеличивается довольно быстро. У одних растений, преимущественно двудольных, такое быстрое увеличение длины продолжается до 20—25 лет, у хвойных — до 60 лет и даже несколько более. Длина члеников сосудов также увеличивается, но так как членики сосудов вообще короткие, прирост длины их менее заметен, чем прирост волокнистых элементов.
После отмеченного периода (25—60 лет) длина прозенхимных элементов уже так сильно и закономерно не меняется, оставаясь, более или менее постоянной с незначительными колебаниями в ту или другую сторону в зависимости от ряда обстоятельств. К числу факторов, влияющих на колебания в длине трахеид и других прозенхимных элементов, прежде всего следует отнести изменения климатических условий и общих условий произрастания древесного растения из года в год.
Длина анатомических элементов на любом уровне стебля древесного растения по направлению от центра к периферии тоже неодинакова. Сначала длина увеличивается до некоторого предела, а потом становится более или менее стационарной. Неодинакова и длина анатомических элементов вдоль по стеблю от основания до вершины. Сначала она постепенно увеличивается, достигая на некоторой высоте максимума, а далее к вершине снова уменьшается.
Итак, длина анатомических элементов проводящей системы стебля как в горизонтальном, так и вертикальном направлениях неодинакова и изменяется с некоторой закономерностью. Несомненно, эти закономерные изменения длины анатомических элементов, установленные для древесины большого числа древесных растений, существуют и у анатомических элементов флоэмы тех же растений, а также у анатомических элементов проводящей системы травянистых растений.
Приведем несколько примеров длины трахеид во вторичной ксилеме древесных растений.
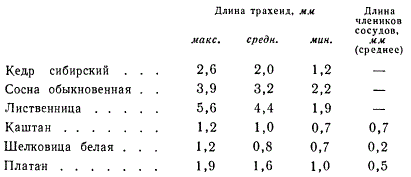
Приведенные цифры показывают, насколько длина одних и тех же анатомических элементов у голосеменных больше, чем у покрытосеменных двудольных. Уменьшение длины трахеидоподобных анатомических элементов во вторичной ксилеме двудольных тесно связано с развитием сосудов. Трахеидные клетки у двудольных становятся все короче и короче, в то время как сосуды приобретают все более и более специализированную структуру. Наиболее специализированным сосудом следует считать сосуд, состоящий из коротких широких члеников с простыми перфорациями, причем боковые стенки их густо усеяны округлыми или ограниченными окаймленными порами.