Принимается, что ствол приматов, в трактуемом нами объеме, берет свое начало от верхнемеловых насекомоядных, относящихся к группе тупай (Tupaioidea), современные представители которых обитают на Яве, Борнео, Суматре, на южной оконечности Малайского полуострова и Филиппинах.
От древних тупай происходят современные тупаи, лемуры, долгопяты и широконосые (американские) обезьяны. Эти группы и их эволюцию мы оставим в стороне. Корнем обезьян старого света считаются эоценовые некролемуры — вымершая группа своеобразных долгопятов, имеющих ряд черт сходства в зубной системе с настоящими обезьянами. Таким образом, древнетретичные долгопяты, повидимому, играли роль корня обезьян старого света.
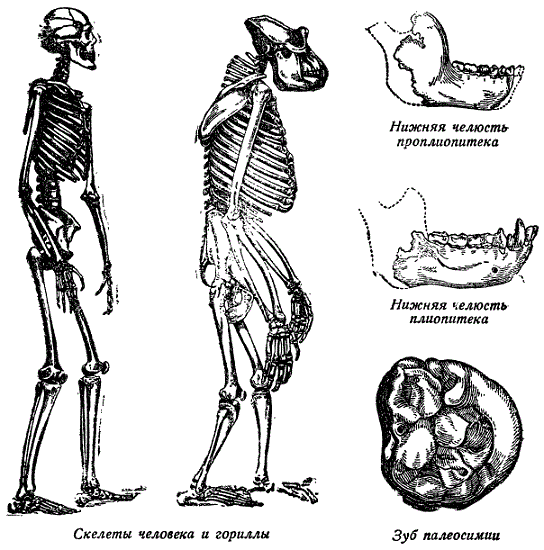
Древнейшей антропоморфной обезьяной считается фраасов парапитек (Parapithecus fraasi), найденный в нижнеолигоценовых слоях Фаюма (Египет). Зубная формула его (i 2с 1 pm 2m 3) соответствует формуле современных антропоморфных обезьян. Клык маленький и, судя по размерам челюсти (36 мм), это была маленькая обезьяна. Ближайшим потомком парапитека считается проплиопитек Геккеля (Propliopithecus Haeckeli), описанный по нижней челюсти из тех же олигоценовых фаюмских слоев. Челюсть этой обезьяны значительно больше, чем у парапитека, причем клык еще невелик и едва выступает над зубным рядом. Челюстная рама, кроме того, значительно массивнее, чем у предыдущей формы. Весь зубной ряд относительно низкий, как и у парапитека.
С последним проплиопитек сближается по форме ложнокоренных зубов и общей примитивности, в частности — относительно слабой диференцировке зубного ряда.
От проплиопитека можно наметить два направления эволюции: одно ведет к плиопитекам, другое к палеосимиям и дриопитекам.
Плиопитек (Pliopithecus) известен по останкам верхних и нижних челюстей. Последние значительно превосходят по размерам челюсти предшествующей обезьяны. Клык возвышается над зубным рядом. Верхние и нижние коренные сходны с коренными современных гиббонов. Плиопитеки, повидимому, достигали их роста и населяли в миоцене леса Средней Европы. Принимается, что плиопитек является предком современных гиббонов.
Проследим теперь второе направление, ведущее к современному орангутану. За основание ветви орангов принимается палеосимия (Palaeosimia), известная по одному коренному зубу. Зуб был найден в Индии, в слоях Сивалика, относящихся к верхнему миоцену. В тех же сиваликских горизонтах были обнаружены фрагменты челюстей сивапитеков (Sivapithecus). Допускается, что сивапитеки (S. sivalensis и S. indicus) были непосредственными предшественниками орангутана, останки которого впервые появляются в плиоцене Восточной Индии, в верхнем сиваликском горизонте.
Эволюция ветви гиббонов и орангутанов пошла по пути специализации к древесному. образу жизни, особенно явственно и типично выраженному у гиббонов. Основной признак гиббонов заключается в относительно небольшой величине тела и его легкости, а с другой стороны, — в интенсификации лазательных функций. Лазательная функция у обезьян приняла особый характер брахиации, при которой передние конечности используются как подвески при перебрасывании по воздуху с одной ветви на другую. Брахиация несомненно давала дендробионту большие преимущества, обеспечивая быстроту перемещения с дерева на дерево и повышая способность к конкуренции в использовании дерева, как кормовой базы. К этому преимуществу у предков гиббона и антропоморфных обезьян прибавилась еще одна черта, состоящая в том, что у всех этих форм постепенно выработалось отвесное положение тела с перенесением центра тяжести на таз и нижние конечности. Это имело своим последствием диференцировку функций передних и задних конечностей. Функции последних подверглись сужению и ослаблению. Напротив, для передних конечностей можно констатировать интенсификацию функции брахиации, при сохранении других функций (хватание предметов, срывание плодов, очистка кожуры с них, постройка гнезда, оборона и т. п.), свойственных передним конечностям обезьян. У гиббонов эти преобразования связаны с другими чертами организации: относительно высоким развитием мозга, обостренностью зрения, необычайно развитым глазомером, с небольшим весом тела. Совокупность всех этих моментов привела к выработке поразительно совершенного типа брахиатора, обладающего невероятной ловкостью, точностью и легкостью движений, что позволило гиббонам успешно конкурировать с другими дендробионтами.
Однако, при небольшом весе тела гиббонов, брахиация привела к признакам узкой специализации, близкой к типу теломорфозов. Это выражается, в частности в образе жизни гиббонов, типичных, довольно узко специализованных дендробионтов, совершенно чуждых хтонобионтности. Поэтому эволюция гиббонов, в конечном счете, не дает выходов к более высокой организации и представляет собою замкнутую, специализованную ветвь, хотя и близкую к антропоморфным.
Выход из условий этой специализации возник на пути увеличения размеров и веса тела. Последствия этого приобретения сказались даже на таком относительно специализованном дендробионте-брахиаторе, каким является орангутан. Быстрота движений, при большом весе тела, снизилась, мощь животного возросла. Вес самцов орангутана, как мы видим, достигает 70—100, а иногда 200 кг. При таком весе понятна осторожность и медлительность движений, которые характерны для оранга.
Брахиация в этом случае не может вести к такой степени специализации, как у гиббонов и приближается к понятию идиодаптаций в широком смысле слова (алломорфоз Шмальгаузена). Менее узкая специализация в направлении брахиаций привела к более высокому развитию. Так, мозг гиббонов примитивнее, чем мозг оранга, похожий на человеческий. Рука оранга менее специализована. Это отражено в ее строении. У гиббонов, например, у яванского, первый палец значительно короче остальных, чем у оранга. Тем не менее и ветвь орангутанов остается боковой по отношению к человеку. Его нишей является дерево. Орангутан — типичный дендробионт, тогда как предки человека развивались, как хтонобионты.
Обратимся теперь к третьему направлению — ветви, ведущей к шимпанзе, горилле и человеку.
Исходной формой для них являются потомки проплиопитека — дриопитеки (Driopithecus).
Все они были крупными обезьянами, достигавшими размеров шимпанзе. Они жили в количестве целого ряда видов в миоцене Европы, Египта, а также известны из нижне-плиоценовых слоев Ю. Азии и Европы.
Большое число видов указывает на биологический прогресс этой группы, а их местонахождения — на широкий родовой ареал. Зубы дриопитеков были примитивнее, менее специализованы, чем у современных африканских человекообразных. Во многих отношениях это — сборная группа, в пределах которой наблюдаются отдельные черты организации современных африканских человекообразных. Так, строение нижнего третьего коренного (м3) у Dr. chiniensis и современного гориллы очень сходны. С другой стороны, зубная система последней сходна с таковой фонтанова дриопитека (Dr. fontani и penjabicus), и возможно, что последний (пенджабский дриопитек) является предком гориллы. Dr. germanicus близок к шимпанзе. Повидимому, Dr. germanicus является все же боковой ветвью и непосредственным предком рода шимпанзе нужно считать миоценового проконсула (Proconsul africanus).
Таковы предки обеих современных африканских человекообразных обезьян. В построении этой схемы филогенетических отношений ведущее значение имеет строение зубов перечисленных видов, их форма, количество бугорков, структура корней и т. д., на чем мы здесь останавливаться не можем.
Что характеризует эволюцию перечисленных форм? Несомненно, что ведущим их признаком являются крупные размеры и переход от чисто древесного к частично наземному существованию.
Способ лазания по деревьям изменяется. Наблюдая за движениями шимпанзе, можно видеть, что он сохранил способность к брахиации. Однако, во-первых, и здесь крупные размеры и вес препятствуют узкой специализации. Во-вторых, эта ветвь человекообразных приобрела, в ходе эволюции крупных дриопитеков, другой способ движения, опять-таки связанный с отвесным, положением тела. Этот способ называется круриацией (crus — нога). При круриации в восхождении на дерево важную роль играют задние конечности, на которые опирается выпрямленное, отвесно ориентированное тело, тогда как руки становятся не подвесками, а органами придерживания за ветви. Движение по способу круриации, в условиях приближенно-вертикального положения тела, получило большое значение. Дерево перестает быть единственной кормовой базой. Таковой становится отчасти почвенный, т. е. нижний ярус лесной растительности (в частности корнеплоды). Таким образом, ниша крупных дриопитеков, несомненно, эволюировала в сторону хтонобионтности. Поэтому уже шимпанзе очень часто встречается на земле, тогда как более тяжелый горилла становится хтонобионтом. Фактически, однако, и на земле обе эти обезьяны передвигаются по способу круриации. Выпрямившись, шимпанзе и горилла всей своей тяжестью опираются на задние конечности, придерживаясь за ветви или землю передними конечностями. Рука не стала свободной. В развитии их ниши ведущую роль все же играет лес. Все антропоморфные по преимуществу — растительноядные, тогда как в эволюции к человеку, как увидим ниже, большую роль сыграла плотоядность (хищничество). Массивные челюсти и зубы связаны с мощным развитием жевательной мускулатуры и — коррелятивно — с мощным развитием гребней, — обстоятельство, сказавшееся на пределах развития черепной коробки. С другой стороны, у антропоморфных нет посылок к развитию речи, их челюстная дуга и язык узкие, ротовая полость не приспособлена к артикуляционной функции, в мозгу не развиты речевые центры, череп не сбалансирован, способность к вертикальному хождению ограничена, конечности специализованы, как конечности дендробионтов. Организация человекообразных обезьян — алломорфного типа, без выходов к адаптациям широкого значения, к ароморфным преобразованиям. Гоминизация (очеловечение) могла быть достигнута на путях менее специализованной и, следовательно, более примитивной организации.
Перейдем теперь к ветви людей (Hominidae). Основным признаком ее является переход к прямостоянию при полном освобождении рук от функции опоры тела. Что же явилось посылкой к этим важным преобразованиям? Мы видели, что отвесное положение тела возникло уже и при брахиации, тем более при круриации. Как указывалось, это привело к перенесению центра тяжести на таз и задние конечности, что связано с рядом изменений в строении этих частей тела. Таз стал шире, крестцово-подвздошные сочленения прочнее. Произошли также изменения в строении задних конечностей, в особенности, развитие куполообразной стопы и параллельное остальным положение большого пальца.
Таким образом, прямохождение имеет некоторые анатомические предпосылки. Однако, ими одними проблема не решается. Вторым условием был переход к полной хтонобионтности. Посылкой к развитию последней был экологический фактор — эволюции ниши, завоевание новой стации — безлесных пространств с большим кругозором и широким горизонтом. Только в этих условиях прямостояние развивается, как биологическая необходимость. Вопрос идет о расширении местообитаний за счет открытых, возможно скалистых пространств.
Ветвь гоминид имеет своим источником тех же дриопитеков, возможно — Dr. Darwini. Она начинается рамапитеком — обезьяной, от которой сохранились фрагменты верхней и нижней челюстей нижнеплиоценового геологического возраста. Характерно, что рама челюсти относительно тонкая и высокая. Ее реконструкция позволяет считать, что челюсть имела форму параболической дуги, приближающейся к сравнительно короткой челюсти последующих гоминид. Рамапитеков, по современным данным, можно считать предками замечательной плейстоценовой африканской обезьяны — австралопитека (Australopithecus), возбуждающего живейший интерес. Это была человекообразная обезьяна, населявшая в указанное выше время открытые пространства и, несомненно, уже владевшая походкой на двух задних ногах. Был найден почти целый череп австралопитека. Этот последний значительно более «человечен», чем у гориллы и шимпанзе. Надглазничный валик выступает слабо, челюсти не столь сильно выдаются вперед, клыки значительно короче, диастема отсутствует, структура жевательной поверхности зубов приближается к человеческой, вся челюсть в целом относительно короткая, образует округлую дугу. Указанный череп относился к молодой особи, что умаляло значение описанных признаков. Но в 1936 году, снова в Ю. Африке, был найден череп взрослого австралопитека, который позволил реконструировать облик этой обезьяны и установить, что она образует переходную форму от рамапитека к гоминидам. Кроме австралопитека были обнаружены и другие прямостоящие обезьяны (плезиантроп, парантроп). Повидимому, в верхнем плиоцене существовало несколько видов прямостоящих антропоморфных обезьян, близких семейству людей (гоминидам).
Судя по костным останкам, обнаруженным в местонахождении австралопитека, можно думать, что в отличие от других, в том числе и современных человекообразных обезьян, австралопитек питался не только растительной, но и животной пищей — явление, имеющее, как увидим дальше, большой теоретический интерес. Австралопитеки и другие прямостоящие обезьяны вымерли в плейстоцене, т. е. на заре четвертичного периода. Однако, за время своего существования они успели широко распространиться в пределах Старого Света, проникнув также в Ю. Азию.
Четвертичный период начинается плейстоценом, характеризующимся значительно более суровым климатом, чем конец третичного времени (плиоцен), и неоднократной сменой холодных ледниковых и теплых межледниковых эпох. Насчитывается четыре ледниковых периода и три межледниковых. В течение плейстоцена происходят существенные изменения в климате, фауне и флоре земли. Нельзя, конечно, рисовать себе ледниковый период, как какую-то катастрофу. Однако, периоды наступления ледников все же существенно отражались на ландшафтах, на флоре и фауне и вызывали глубокие сукцессии в биоценозах. Третичная фауна частью отступает на юг, а в значительной степени вымирает под влиянием новых форм конкуренции в условиях более острой борьбы за существование. Вымерли и все третичные обезьяны, в том числе и дриопитеки, австралопитек и его боковые ветви. Однако, прямостоящие обезьяны вымерли не бесследно, и это новое направление эволюции приматов сохранилось. Объясняется этот факт тем, что у прямостоящих обезьян мы видим возникновение ряда ароморфозов и, с другой стороны, — отсутствие признаков узкой специализации. В пользу последнего предположения говорит, в частности, строение челюстного аппарата, свойственное, например, австралопитеку. Мы видели, что его челюсть относительно короткая, клыки не выступают, диастемы нет, сагиттальный гребень отсутствует, надглазничный валик относительно слабо выражен. Эти черты — примитивнее, свидетельствуют о меньшей специализации, чем у современных человекообразных обезьян. С другой стороны, прямостоящие обезьяны обладали рядом важных ароморфозов. Огромную роль в их развитии сыграло прямохождение. Последнее создавало ряд существенных преимуществ экологического характера. Сюда относится расширение горизонта, значительно более дальнее и широкое обозрение местности, а следовательно, гораздо большая возможность своевременной регистрации опасности или, напротив, подготовки к нападению на других животных, наконец, — общей ориентировки в окружающей местности. Однако, еще более важно было освобождение передней конечности от ее примитивной функции, как органа передвижения. Свободная рука, уже на заре эволюции прямостоящих обезьян, приобрела огромное значение, как орган защиты и нападения. Редукция зубного аппарата у австралопитека показывает, что зубы, как орган зашиты, теряют ведущую роль. С другой стороны, тот факт, что австоалопитек употреблял в пищу мясо, позволяет думать, что ему было свойственно хищничество. Констатация последнего делает весьма вероятным предположение, что уже у австралопитека рука стала совершенно особым органом защиты и нападения, заменившим редуцирующийся зубной аппарат, — а именно, органом дальнего действия, усиленным при помощи соответствующего вооружения. Удар обломком ветви, а тем более метание камней на расстояние приобрели большое значение, как орган защиты и агрессии. В отношениях прямостоящих обезьян с остальным животным миром, прямостояние, вероятно, имело также известное значение, как постоянно действующая «поза угрозы». Мы знаем, что многие животные, в момент раздражения, поднимаются на задние ноги и что эта поза имеет значение позы угрозы. Напомним, что она очень характерна для приведенного в состояние ярости самца гориллы и что, с другой стороны, вертикальная фигура человека вызывает у многих животных реакцию в форме бегства. Едва ли можно сомневаться в том, что прямостоящие обезьяны сохранили свойственную многим питекоидам черту, а именно — стадность, в условиях которой сообща действовали несколько индивидуумов, что, конечно, делало прямостоящих обезьян опасным соперником. Их защитные и агрессивные реакции становились особенно действенными вследствие другого, важнейшего, фактора — высокого развития головного мозга. По всем этим соображениям можно принять, что хотя австралопитек и другие прямостоящие обезьяны плейстоцена и вымерли, но это вымирание было лишь следствием конкуренции с другими, более высоко организованными прямостоящими приматами, продвинувшимися в своем филогенетическом развитии еще дальше по пути гоминизации.