Проблема факторов эволюции составляет центральную проблему системы дарвинизма. Уже указывалось, что ведущими факторами эволюции являются: изменчивость, наследственность и отбор.
Обратимся к рассмотрению изменчивости. Учение об изменчивости слагается из следующих подчиненных проблем:
- Определение понятия об изменчивости.
- Формы изменчивости.
- Причины изменчивости.
- Значение различных форм изменчивости в эволюционном процессе.
Вопросы изменчивости трактуются не только в системе дарвинизма, но также в пределах других биологических наук. Любое биологическое явление может быть освещено с различных сторон. Задачей дарвинизма, как науки, должно быть изучение изменчивости, как первичного фактора эволюционного процесса. Решение этой задачи отчасти связано с точным определением явления изменчивости. В основу должны быть положены работы Ч. Дарвина. Большое значение для выработки дарвинистского определения изменчивости имеют также работы К. Тимирязева.
Определение понятия изменчивости
Дарвин уделял изменчивости огромное внимание. Ей посвящена особая глава «Происхождения видов» и ряд глав другой его работы «Изменение животных и растений в состоянии одомашнения». Анализ дарвиновской постановки проблемы изменчивости выясняет следующие положения.
Во-первых, Дарвин рассматривал изменчивость, как процесс, который проявляется в результате полового и бесполового размножения. Во-вторых, Дарвин стремился показать, что изменчивость, сама по себе, не есть эволюционный процесс, т. е. не адекватна ему, оставаясь лишь элементарным источником эволюционного процесса, в частности процесса видообразования. Эти представления легли также в основу взглядов К. А. Тимирязева. В последарвиновскую эпоху изменчивость изучалась многими исследователями в этом же освещении. Однако, в начале XX века проблема изменчивости была вовлечена в русло антидарвинистических представлений. Это повлияло на общую оценку явления изменчивости. Учение об изменчивости было выделено из системы дарвинизма и вошло в состав генетики. Органические связи между учением об изменчивости и системой дарвинизма были, таким образом, в значительной степени утрачены, а сам дарвинизм стал трактоваться, как прошлое науки.
Так, Иогансен (1903), обосновывая положение учения об изменчивости в системе генетики, пришел к выводу, что можно различать три группы явлений изменчивости:
1) Различия внутри наиболее узких систематических групп, т. е. внутри «чистых рас», входящих в состав вида; 2) различия видовые, характеризующие виды; 3) различия, наблюдаемые у бастардов, т. е. у форм, полученных в результате скрещивания.
По Иогансену, генетику интересуют только первая и третья группа изменений. Вторая группа изменений — предмет исследования систематиков. О связях проблемы изменчивости с эволюционной теорией, а тем более с дарвинизмом, в этой классификационной схеме — ничего не говорится. Иогансен указывает также, что учение о наследственности (куда включается и учение об изменчивости) лучше всего изучать независимо от эволюционного учения, тогда как последнее немыслимо без первого.
Из сказанного вытекают следующие выводы. Во-первых, изменчивость можно трактовать, как явление, как бы не имеющее обязательных связей с процессом эволюции. Во-вторых, в соответствии с этим положением, изменчивость можно рассматривать, как явление различий. Такое понимание изменчивости получило широкое распространение и вошло в литературу, в том числе учебную. Так, Филипченко (1915) предложил следующее определение изменчивости: «мы понимаем под изменчивостью наличие различий как между особями, так и между группами особей, относящимися к одному виду».
Им было также подчеркнуто, что изменчивость можно изучать двояко: как состояние (наличие различий) и как процесс. На возможность этих двух аспектов указал еще Дженнингс (1908). Филипченко считает, что первое понимание изменчивости статично, тогда как второе рассматривает динамику изменчивости.
Изучение «изменчивости, как статики», получило широкое распространение, и в ряде случаев оно удовлетворяло исследователей, изучавших изменчивость вне проблем эволюции. К. А. Тимирязев тотчас обратил на это внимание и указал, что «нередко смешивают изменчивость с простым фактом наличности различий». По Тимирязеву, под изменчивостью «должно разуметь… превращения органических существ, возникающие во времени». Таким образом, Тимирязев рассматривал изменчивость, как процесс. Именно это понимание изменчивости и должно быть положено в основу дарвинистской интерпретации ее.
Вторым вопросом, который был поднят Тимирязевым, является проблема содержания процесса изменчивости. Им было указано, что речь идет о возникновении «совершенно новых особенностей строения или отправления» организмов. Наконец, по Тимирязеву, в определение изменчивости должно быть включено представление о таких новых изменениях, которые означают «отклонения от видового типа».
Синтезируя сказанное выше, мы будем придерживаться следующего определения изменчивости: изменчивость есть процесс возникновения конкретных новых особенностей, представляющих отклонения от видового типа и ведущих к развитию различий между особями.
Это определение ближе к задачам дарвинизма, так как оно трактует изменчивость, как материал для эволюционного процесса. С другой стороны, оно не закрывает и не может закрыть возможности и необходимости изучать различия, так как исследование последних остается главным источником наших знаний о самом процессе изменчивости и ее результатах. Однако не надо забывать, что явление различий есть не изменчивость, а ее результат.
Таким образом, различия безусловно нужно изучать. Однако, нужно помнить, что они возникли и возникают и, что то состояние различий, которое исследователь регистрирует, есть известный этап процесса изменчивости, засеченный в момент наблюдения.
Вопреки соответствующим представлениям Дженнингса, Филипченко и других исследователей, явление различий не может быть противопоставлено, как статика динамике. Напротив, явление различий есть выражение динамики изменчивости, ее материальное осуществление, без которого эмпирическое познание изменчивости было бы невозможно.
Формы изменчивости
Синтезируя данные современной ему науки, Дарвин предложил различать несколько форм изменчивости.
Дарвин различал, прежде всего, изменчивость наследственную и ненаследственную. Эта диференцировка, как легко видеть, касается содержания явления. Кроме того, Дарвин различал также процесс изменчивости по его формам. Как указывалось, он дал характеристику следующих форм изменчивости: определенная, неопределенная, коррелятивная и изменчивость вследствие скрещивания.
Под влиянием работ Дарвина изменчивость подверглась всестороннему изучению. В ходе этих работ частично была предложена новая терминология форм изменчивости, которая и удержалась в науке. Она применяется и в этом курсе дарвинизма. Однако согласование терминологии Дарвина с современной совершенно необходимо. Терминология Дарвина была несправедливо забыта, и это повлекло за собою некоторые недоразумения и путаницу, которых нужно избежать.
В таблице дается принятая классификационная схема форм изменчивости, сопоставленная с терминологией Дарвина.
| Современная терминология | Терминология Дарвина |
| 1. Мутация | 1. Наследственная неопределенная индивидуальная (единичная) изменчивость. |
| 2. Модификация | 2. Ненаследственные индивидуальные или массовые определенные изменения. |
| 3. Корреляция | 3. Коррелятивная или соотносительная (зависимая) изменчивость. |
| 4. Комбинативная изменчивость | 4. Изменчивость, возникающая вследствие скрещивания. |
Таким образом, неверно ставить знак равенства между мутацией и дарвиновской неопределенной изменчивостью, что иногда делается. Можно лишь поставить знак равенства между термином «мутация» и термином наследственная неопределенная изменчивость. Столь же неверна попытка приравнивать термин «модификация» к термину Дарвина «определенная изменчивость». В этом случае необходимо оговорить, что модификация есть ненаследственная определенная изменчивость — индивидуальная или массовая.
Некоторые авторы внесли путаницу в понятие индивидуальной изменчивости. Начало проблеме индивидуальной изменчивости в ее антидарвинистском понимании было положено Гуго де-Фризом, который предложил различать две формы изменчивости: индивидуальную, или флюктуирующую (флюктуации) и видообразующую. Первая не наследственна, т. е. не сопровождается изменениями наследственного основания организма и соответствует (терминологически) модификациям. Вторая, наоборот, наследственна и терминологически соответствует мутациям. Очевидно, только наследственная изменчивость может обеспечить возникновение нового наследственного основания.
Так как Дарвин придавал основное значение индивидуальной изменчивости, то де-Фриз сделал отсюда неправильный вывод, что Дарвин основывал свою теорию эволюции на ненаследственной, т. е. флюктуирующей, индивидуальной изменчивости, и что, следовательно, его теория построена на неверной посылке. По де-Фризу, Дарвин строил теорию накопляющего действия отбора на индивидуальных изменениях (флюктуации), которые, как ненаследственные, не могут закрепляться в потомстве, а следовательно, и накопляться. Де-Фриз упустил, что Дарвин имел в виду наследственную индивидуальную изменчивость. Ошибка де-Фриза не была замечена и его представления получили широкое распространение, пока их ложность не была указана Плате (1910).
Рассмотрев классификационную схему изменчивости, перейдем к систематическому изучению форм ее. Предварительно, однако, остановимся на некоторых важных терминологических понятиях, без которых излагать учение об изменчивости трудно.
Основные терминологические понятия
Современная наука выработала ряд понятий, в высокой степени облегчающих понимание процессов изменчивости.
А. Генотип и фенотип. Термины эти были предложены Иогансеном (1903). Фенотип Иогансен определяет следующим образом: «Фенотип любого индивидуума есть сущность всех его внешне проявляющихся свойств». Таким образом, фенотип каждого индивидуума определяется его морфологическими и физиологическими признаками. Они и составляют его фенотип. Следует указать, что фенотип индивидуума развивается в процессе онтогенеза и, следовательно, изменяется. Фенотип взрослой особи не стабилизируется. Изменения фенотипа продолжаются до конца жизни индивидуума. Таким образом, смерть является естественным завершением развития фенотипа. Нужно, однако, помнить, что фенотип каждой особи определяется не только ее индивидуальными чертами. Уже указывалось, что любая особь обладает, кроме того, и более общими чертами, в частности — видовыми. Если в ходе онтогенеза развиваются индивидуальные фенотипические черты особи, то параллельно им развиваются и ее видовые черты. В практике это положение подтверждается тем, что, например, описание новых видов возможно сплошь и рядом только в том случае, если исследователь имеет дело со взрослыми особями.
Одним из факторов формирования фенотипа особи является наследственное основание ее, или ее генотип (Иогансен). Как правило, особи, имеющие разные генотипы, характеризуются различными фенотипами. Изменение генотипа влечет за собою изменение фенотипа — направление, характер и форму его развития. При современном состоянии наших знаний и в свете материалистической диалектики можно утверждать, что в природе не существует двух абсолютно одинаковых генотипов. Отчасти по этой причине не существует и двух абсолютно одинаковых фенотипов. Изложенные данные показывают, что эндогенные факторы играют большую роль в осуществлении фенотипа.
Вторым компонентом фенотипа являются внешние или экзогенные факторы, участвующие в формировании организма. Изменяя генотип, внешние факторы косвенно воздействуют и на его фенотипическое осуществление. В то же время внешние факторы воздействуют на фенотип и непосредственно. Дальше эти отношения разбираются подробнее. Следствием кратко описанных отношений должно быть огромное гено — и фенотипическое разнообразие живых форм, входящих в состав одного и того же вида. Население или индивидуальный состав вида неизбежно оказывается генотипически и фенотипически неоднородным, разнокачественным. Эта внутривидовая система генотипически и фенотипически неоднородных индивидуумов и составляет популяцию вида.
Б. Фенотипическая изменчивость. Из сказанного выше видно, что если мы, например, в полевых условиях, наблюдаем явления неопределенной (единичной) изменчивости, то мы не всегда можем сказать, с какой изменчивостью мы имеем дело — ненаследственной или наследственной. В самом деле, быть может, данное единичное изменение есть только модификация, т. е. ненаследственное изменение, или, наоборот — мутация, т. е. изменение самой наследственной основы. Решается этот вопрос путем эксперимента, в частности — испытания потомства (в особенности второго и последующих поколений). Если в потомстве новое индивидуальное изменение фенотипически проявится, и притом хотя бы и в несколько измененных условиях, то такое изменение, очевидно, наследственно (мутация). Если этого нет, и изменение не только в первом, но и во втором и последующих поколениях не проявится, а наоборот — исчезнет, то правильнее считать его ненаследственным (модификация).
Итак, в условиях простых наблюдений мы часто не можем заранее определить, имеем ли мы дело с индивидуальной модификацией, или с мутацией.
Однако и в том и в другом случае изменение налицо, так как оно проявилось в конкретных морфо-физиологических, видимых или вообще познаваемых, фенотипических изменениях. Следовательно, например, в полевой обстановке следует говорить в самой общей форме о фенотипической изменчивости. Более же точный экспериментальный анализ позволяет вскрыть истинное содержание ее. Индивидуальная неопределенная изменчивость, о которой писал Дарвин, и есть фенотипическая изменчивость. Когда Дарвин говорит, что индивидуальная изменчивость «как известно, часто наследственна», то это положение и означает, в переводе на современную терминологию, что фенотипическая изменчивость, как известно, часто носит мутационный характер.
Очевидно, фенотипическая изменчивость есть общее выражение факта изменчивости, включающей и модификационную и мутационную изменчивость.
В. Мутации, модификации и признаки. Итак, исследователь имеет дело, прежде всего, с фенотипом, т. е. с конкретными морфологическими и физиологическими признаками (цвет, запах, вкус, форма, пропорции, размеры, число частей и т. д.).
Возникает вопрос о взаимоотношениях между понятиями: признак, мутация, модификация.
Ответ на этот вопрос вытекает из нашего определения изменчивости. Как мы видели, изменчивость есть процесс возникновения новых особенностей. Термины «модификация» и «мутация» обозначают самый процесс или ход изменения, его становление и развитие. Признак же есть не модификация или мутация, а фенотипический, т. е. видимый результат модификационного или мутационного процесса.
Поэтому нужно строго различать следующие понятия: изменения, т. е. модификации и мутации и результаты изменений — новые признаки. Носителей новых признаков можно назвать соответственно модификантами и мутантами.
Остается решить последний вопрос — о наследственности признаков. Если следовать изложенной выше схеме представлений, то необходимо признать, что признаки, сами по себе, не наследственны. Можно лишь говорить о наследственности изменений. Что же касается новых признаков, возникающих в результате изменений, то они являются лишь фенотипическим выражением последних, так как признаки зависимы, с одной стороны, от «внутренних» особенностей организма, а с другой, — от условий жизни.
Это доказано совершенно твердо, на огромном количестве фактов. Приведем некоторые из них. Известно, что породные признаки культурных животных проявляются только в условиях соответственного кормления и общего благоприятствующего содержания. При плохом кормлении не проявятся и типичные наружные признаки породы, ее экстерьер. Особенно убедительны экспериментальные данные, касающиеся признаков, имеющих геологическую давность, которые, казалось бы, имели время наследственно закрепиться. Например, всем билатеральным животным свойственно наличие правого и левого глаза. Эта особенность возникла в геологическом прошлом и существует поныне. Если подвергнуть яйца рыб или амфибий, например, действию хлористого магния, то развиваются формы, у которых посредине головы имеется лишь один глаз (так называемая циклопия). Следовательно, наличие двух глаз, само по себе, ненаследственно. Этот признак возникает при нормальных условиях. Однако при изменении их (например, воздействие хлористого магния) признак не возникает, наоборот, возникает новая особенность — циклопия. Подобные явления имеют всеобщий характер. Признаки могут быть изменены переменой условий жизни. С этим фактом мы еще столкнемся неоднократно в дальнейшем.
Подчеркнем однако уже здесь, что изложенная постановка вопроса открывает широкие возможности управления отдельными признаками и фенотипом в целом.
Если это верно, то возникает вопрос: чем же отличаются модификации от мутаций? И те и другие конкретизируются в определенных фенотипических чертах и, следовательно, выражаются сходным образом. Различия между ними таковы. Модификационное фенотипическое изменение есть реакция одного и того же генотипа на разные условия среды. В разных условиях среды один и тот же генотип осуществляется в разных фенотипах.
Очень наглядно этот факт был показан в опытах Боннье (1895), который расчленял одно и то же растение на две продольные половины. Одна половина высаживалась в условиях горного климата, другая— долинного. В данном случае однородность генотипического материала оставалась несомненной. Тем не менее развившиеся особи — горная и долинная —резко отличались друг от друга фенотипически. Оба модификанта были фенотипическим результатом разных условий среды, воздействовавших на один и тот же генотип.
Обратимся теперь к мутационным изменениям. Эти последние являются реакциями измененного генотипа.
Два разных генотипа в одинаковых условиях среды обычно ведут к осуществлению разных фенотипов.
Для пояснения воспользуемся сперва воображаемым примером.
Под влиянием пониженной и повышенной влажности кожа прыткой ящерицы (Lacerta agilis) темнеет. Предположим, что в популяции этой ящерицы появилась особь, которая реагирует на пониженную температуру и повышенную влажность не потемнением кожи, а посветлением ее. Такой случай означал бы, что эта особь есть мутант, т. е. фенотипический результат мутации генотипа. Что здесь изменилось? Очевидно, мы имеем дело с новой формой ответа, или новой нормой реакции на прежнее воздействие условий среды. Следовательно, каждому генотипу свойственна специфическая норма реакции. Мутация и выражается в наследственном изменении нормы реакций генотипа на воздействие факторов среды. Другими словами, мы имеем дело с новым генотипом, т. е. с новым наследственным основанием организма.
Таким образом, если в потомстве, среди типичных для данных условий среды фенотипов, внезапно появляется единичный новый фенотип, всегда можно предполагать, что исследователь имеет дело с мутантом. Это предположение приобретает более высокую степень вероятности, если при тех же условиях среды признаки предполагаемого мутанта появятся и в потомстве.
Рассмотрев перечисленные выше терминологические понятия, обратимся к более подробному изучению форм изменчивости.
Наследственные неопределенные (единичные) изменения, или мутации
Термин «мутация» был введен в науку де-Фризом (1900,1901), хотя применялся и раньше (Адансон). Русский исследователь С. Коржинский (1899) собрал большое число данных о мутациях, обозначая их, по примеру Келликера (1864), термином гетерогенезис. Де-Фриз понимал под мутациями такие качественные изменения наследственного основания организма, которые внезапно, скачкообразно создают новые биологические формы и даже виды. Де-Фриз имел в виду защитить положение, согласно которому новые формы создаются не отбором, а самим мутационным процессом. С его точки зрения роль отбора не творческая. Он лишь уничтожает одни готовые виды и сохраняет другие.
Это неверное представление о мутациях было использовано антидарвинистами, но подверглось заслуженной критике со стороны некоторых дарвинистов, в том числе Тимирязева. В ходе исследований де-фризовское понимание мутации было отброшено.
В системе дарвинизма под мутациями понимаются наследственные изменения генотипа, выраженные в изменении нормы реагирования на условия среды, вследствие чего среди особей обычного для данного вида фенотипа, при тех же условиях, появляются, как правило, единичные новые фенотипы, возникающие (при тех же условиях) и в последующих поколениях. Новая реакция на прежние условия среды выражается, следовательно, в форме приобретения новых признаков.
Такое понимание мутаций соответствует дарвиновскому представлению о наследственной единичной неопределенной изменчивости. В потомстве типичных родителей среди массы столь же типичных форм появляются отдельные особи с новыми признаками, или мутанты. При этом вновь возникшие особенности передаются потомству, поскольку передалась по наследству измененная норма реагирования на те же условия среды.
А. Распространение мутаций в природе. Многочисленные наблюдения показали, что мутации свойственны как растениям, гак и животным, распространяясь на все системы органов.
У растений известны мутации размеров (карликовость, или нанизм, и гигантизм), формы растительных индивидуумов, мутации покровных тканей, например, исчезновение колючек, мутации строения листьев и цветов, окраски цветов, расположения их на цветоножке, мутации плодов и т. п.

Мутации у растений. 1 — 3 — мутанты львиного зева. Из Филипченко, 4 — мутации у чистотела: нормальная форма, справа — мутант (Из Богданова)
Из перечисленных форм мутационных изменений рассмотрим здесь лишь некоторые примеры. Несомненный мутационный характер носит явление пестролистности, а также возникновение красных листьев. Описаны пестролистный клён, хмель, герани, стручковый перец, гортензия, энотера, кукуруза, тростник и т. д. К мутационным формам относятся краснолистные: кровавый бук, пурпуровый барбарис, орешник, ясень, дуб и т. д.
Из мутационных изменений цветов упомянем о явлении махровости, выражающейся в частичном или полном превращении тычинок в лепестки. Процесс влечет за собою ограниченное или полное бесплодие. Примеры: махровые астры, цикламены, петунии, персик, яблони, терновник, розы и др.
Из мутаций расположения цветов, остановимся на явлении пелории у львиного зева. Цветки этого растения принадлежат к зигоморфному типу (с двубоковой или билатеральной симметрией). Наблюдаются, однако, мутанты, у которых появляется верхушечный цветок, построенный по типу актиноморфного цветка (с лучистой симметрией в расположении частей). Соцветие с таким верхушечным актиноморфным цветком и называется пелорическим. Для многих форм (например, наперстянки) пелория типична. Для львиного зева она не типична, и пелорические соцветия у этого растения носят мутационный характер. Многолетние исследования Баура (1924) показали возникновение ряда других мутаций форм цветка у львиного зева.
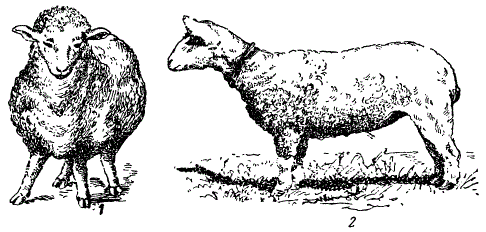
Мутации у животных. 1 — анконская овца, 2 — коротконогая овца, род. в Норвегии (1934) и напоминающая анконскую. (По разным авторам)
Б. Почковые мутации. Многие из описанных выше мутаций возникают не через половое воспроизведение, а вегетативным путем, т. е. в развивающихся почках, следовательно, на ветвях развитого растения.
Большое число данных о почковых мутациях было собрано Дарвином («Изменения животных и растений в состоянии одомашнения»). Сюда относятся, например, появление ветки, несущей красные сливы, на дереве сорокалетней желтой сливы; развитие похожих на персики плодов на ветвях персикового и махрового миндаля; образование позднеспелых персиков на «играющей ветке» менее позднеспелого сорта и, наоборот, раннеспелой формы на нем же; возникновение на ветке вишневого дерева позднеспелых удлиненных плодов; изменение окраски ягод на ветке крыжовника и т. п. В новейшее время данные Дарвина были подтверждены и пополнены. Нередки почковые мутации у винограда, причем, на ветвях определенного сорта внезапно появляются листья или плоды с новыми особенностями. Так, путем почковых Мутаций возникли: полосатость ягод, увеличение размеров грозди, изменение окраски плодов и листьев, пестролистностъ и т. п.
Большое число мутаций описано также для животных.
В. Хроматические мутации, или мутации окраски кожных покровов и дериватов кожи, — одно из наиболее известных явлений.
Распространенной формой хроматических мутаций нужно считать явления меланизма и альбинизма.
Оба типа упомянутых хроматических мутаций наблюдаются у насекомых, рыб, амфибий, птиц и млекопитающих. Таковы: меланистическая форма березовой пяденицы Amphidasis betularia, известная под названием doubledayaria, меланистнческие формы монашенки Porthetria (Liparis) monacha, крыжовенной пяденицы Abraxas glossulariata; альбинистические формы дуболистного шелкопряда Gastropacha quercifolia, аксолотлей, птиц (воробьи, вороны, галки, некоторые дневные хищники, тетерева и т. д.), млекопитающих (мыши, крысы, кролики, лисицы, волки и т. п.).
Хромисты. Альбинизм и меланизм являются лишь крайними случаями хроматических мутаций. Между ними наблюдаются различные другие цветовые морфы. Например, у европейского крота (Talpa europaea) наблюдаются самые различные окраски — от полного альбинизма до черной, причем к последней ведут различнейшие промежуточные окраски — от бледнопалевых до серых и коричневых.

Хромисты тетерева. 1 — albroentris, 2 — brunnea, 3 — andalusica, 4— cholibdea, 5 — alba, 6 — splendees (нормальная окраска). (По Котсу)
Это явление наблюдается и у других форм. Такие цветовые разности получили название хромистое. Была сделана попытка связать хромизм крота с определенными свойствами почвы. Однако аналогичные же хромисты известны для форм, где подобная связь не может быть установлена, например, для многих птиц. Примером могут служить хромисты, описанные Котсом (1937) среди тетеревов. Так, для самцов были установлены 9 типов хромистов, в том числе резко уклоняющиеся по своей окраске от видового типа, например, varietas fumosa с однообразно дымчатым оперением, brunnea землисто-ржаво-охристой окраски, andalusica с окраской типа сизых андалузских кур, chalybdea с белесовато-пепельным оперением и т. п. Большее число хромистов (19) описано Котсом для самок.
Установить какую-либо прямую связь между этими хромистами и, например, приматом — трудно. Окраска типа fumosa относится к особям из Томска, Енисейска, Твери и Вологды. Тетерка типа marginata (землисто-ржавчатая спинка, усыпанная мелкими белесоватыми пестринами) известна Котсу из Скандинавии и Казани. Тетерев альбинос (тип alba) представлен двумя экземплярами: один уральский, другой петропавловский. Отмеченная черта характерна для наследственной неопределенной изменчивости: ее возникновение оказывается не связанным с определенным географическим пунктом, и одна и та же мутационная форма может наблюдаться в различных климатических условиях.
Наряду с изменениями окраски кожных покровов наблюдаются мутационные изменения, связанные с редукцией их, или, наоборот, сильным развитием. Так, у многих млекопитающих наблюдается наследственная бесшерстность, очень мощное развитие волосяного покрова, развитие курчавости и т. п.
Разумеется, мутационный процесс распространяется и на другие признаки. Таковы широко распространенные мутации конечностей и в частности числа пальцев, мутационная бесхвостость (куцехвостые кошки и собаки). К мутациям нужно также отнести кот роткопалость у человека. Описан случай мутационного отсутствия у человека ключиц и т. п.
Из других примеров мутаций напомним многообразные мутации у дрозофилы: изменения крыльев, окраски и числа фасеток глаз, формы брюшка и т. п. Строгие генетические исследования показали, что все эти мутации наследственны.
Ступенчатые мутации. Существование серий незначительных (ступенчатых) мутаций доказано на хорошо изученных объектах, например, на плодовых мухах Drosophila. Так, у этих мух мутирует число фасеток в глазах. На рисунке показан нормально фасетированный глаз, рядом с ним — так называемый лентовидный глаз (мутация Bar-лента) и сверхлёнтовидный глаз (Ultrabar). Эти изменения формы глаз наследственны, и их ряд завершается мутацией, выраженной в полном отсутствии фасеток, т. е. полной слепотой. Другим примером ступенчатых мутаций могут служить мутационные изменения крыльев. Бескрылость связана с полным развитием крыла рядом переходных форм мутаций («зачатковые крылья», «плавничковые», «нерасправленные», «обрубочные» и т. п.).

Мутации у дрозофилы. 4 — 5 нормальное брюшко самца и самки, 6 — 7 — мутационные изменения брюшка. Вверху: мутации глаза: 1 — норма, 2 — bar, 3 — ultrabar.
Г. Частота мутаций. Проблема мелких мутаций. Перечисленные выше примеры показывают, что мутации широко распространены в природе. Они наблюдаются во всех системах органов и, повидимому, у всех живых форм.
По мере исследования существенно изменился взгляд на мутационный процесс. Если первоначально к нему относили резкие, явственно заметные наследственные изменения, то в последнее время были накоплены данные о появлении многочисленных мелких мутаций. Так, исследования Баура над мутациями львиного зева (Antirrhinum majus) вскрыли картину большой их частоты и притом именно мелких мутаций. Баур обнаружил, что мутанты могут отличаться от исходной формы весьма незначительно. Малые мутанты, по данным Баура, «по крайней мере столь же часты, но, вероятно, значительно более часты, чем бросающиеся в глаза мутанты». Баур констатировал, что частота мутирования у Antirrhinum majus достигает 10%. Значит, из каждых 100 гамет десять мутируют. Однако, прибавляет он, это число, в действительности, следует повысить, причем, по его данным, мелкие мутации «распространяются на все особенности растения». У плодовых мух частота мутации достигает 40%, и они распространяются на самые различные признаки — окраску, структуру, величину и форму тела, строение антенн, форму, величину и жилкование крыла, число щетинок на теле, окраску и форму глаз и т. п.
Многие из этих мутаций носят характер мелких, фенотипически едва отличимых от обычных форм. Число мутаций в значительной степени определяется степенью изученности.
Так, в 1922 г. у яблонь было известно около 20 почковых мутаций, а к 1937 г. — свыше 250. Упоминавшиеся выше исследования Баура, а также Тимофеева-Рессовского (1935), Керкиса (1938) и других авторов показали наличие очень большого числа мелких мутаций физиологического характера, едва отраженных в морфологических признаках.
Эти данные подтверждают идею Дарвина, что в эволюции главную роль играют мелкие наследственные неопределенные изменения.
Ненаследственные индивидуальные и массовые (групповые) изменения (модификации)
Термин «модификация» был предложен Иогансеном. В широком смысле слова под модификациями следует понимать ненаследственные изменения, возникшие под влиянием факторов абиотической и биотической среды. К первым принадлежат: температура, влажность, свет, химические свойства воды и почвы, механически действующие факторы (давление, ветер и т. д.), ко вторым пища, а также прямое и косвенное воздействие организмов.
Все эти факторы вызывают ненаследственные фенотипические изменения более или менее глубокого характера.
В естественной обстановке на организм влияют, разумеется, не отдельные факторы, а их совокупность. Однако, некоторые факторы среды имеют ведущее значение. Хотя соответственное значение факторов среды определяется в конечном счете генотипическими свойствами конкретного организма, его физическим состоянием и стадиями развития, все же можно утверждать, что главнейшее модифицирующее значение имеют температура, степень влажности и свет, а для водных; организмов— солевой состав воды.
Температура определяет очень многообразные модификационные изменения. Так, под влиянием различной температуры цветы китайской примулы (Primula sinensis) приобретают различный цвет. При 30—35° развиваются белые цветы Р. sinensis alba, при 15—20° — красные цветы. Р. s. rubra. Листья одуванчика Taraxacum) в период низких температур (4—6°) развиваются в форме глубоко вырезанных пластинок. С наступлением более теплого времени на том же растении развиваются не столь глубоко вырезанные листовые пластинки, а при относительно высоких температурах (15—18°) появляются цельнокрайние листья. Подобные же явления наблюдаются у животных. Так, у бабочек было установлено изменение окраски крыльев под влиянием температуры. Например, у ванесс повышение температуры вызывает усиление красных и желтых тонов. Заметные изменения претерпевает под влиянием температуры окраска кожи земноводных и пресмыкающихся. У травяной лягушки (Rana temporaria) понижение температуры сопровождается посветлением кожи, повышение температуры — потемнением. Темные экземпляры прудовой лягушки при повышении температуры до 20—25° заметно светлеют. То же самое наблюдаются и у саламандр. Наоборот, у других форм, например, у стенной ящерицы (Lacerta muralis) наблюдается потемнение кожи при высокой температуре (37°) и посветление при низкой. Влияя на окраску кожи животных, температура воздействует и на дериваты кожи. Окраска кожи млекопитающих и волосяного покрова также в ряде случаев связана с температурным воздействием. Ильин (1927) показал это на горностаевых кроликах. Удаление шерсти у этих животных с последующим содержанием их на холоду вызывало развитие на выбритых местах черного пигмента и последующего роста черных волос. Известно, что волосяной покров млекопитающих под влиянием пониженной температуры достигает более пышного развития. Этим в известной мере объясняется отмеченный еще Бэром (1845) факт, что мех пушных зверей более сильно развит по направлению к северо-востоку. Температура влияет также на развитие форм тела животных и его придатков. Сёмнер (1909) показал, что воспитание новорожденных мышей в тепле приводит к более слабому развитию шерсти, к удлинению ушей и хвоста. Аналогичные данные подучил Пржибрам (1909) в опытах с крысами. Было показано, что при 30—35° рост крысят идет медленно, а масса тела выросших крыс оказывается меньше, чем у воспитанных на холоду, что в общем соответствует правилу Бергмана. У животных с переменной температурой тела (холоднокровных) наблюдаются обратные отношения.
Под влиянием фактора влажности удивительные превращения наблюдаются у растений. Подводные листья стрелолиста Sagittaria sagittaefolia имеют вытянутолентовидную форму, надводные — на том же экземпляре растения — типичную стреловидную. У болотного лютика, как указывалось ранее, те же отношения вызывают резкие изменения в строении листовой пластинки.
Еще Константен вызвал у болотного лютика явление гетерофилии. Часть листа, погруженная в воду, получила перистую форму, надводная его половина сохранила цельнокрайность.
Лотелье (1893) воздействием влажности вызвал у колючих растений образование листьев вместо колючек. Подобные превращения претерпевает, например, барбарис.
У животных фактор влажности также вызывает явственные изменения. Прежде всего влажность влияет на окраску. Сухость вызывает у лягушек посветление, повышение влажности стимулирует потемнение кожных покровов. Под влиянием влажности, после каждой линьки, у многих птиц наблюдается потемнение рисунка пера.
Понижение влажности действует в обратном направлении, вызывая посветление перьев и волосяного покрова. Аналогичные явлении наблюдал Формозов (1929) у млекопитающих Северной Монголии. По крайней мере одни и те же формы в сухом и влажном климате имеют разную окраску шерсти (посветление в условиях сухости).
Не вдаваясь в подробности, можно утверждать, что факторы влажности и температуры вызывают многообразные хроматические изменения (изменения окраски), оказывая в то же время формообразующее воздействие на организмы.
Свет также вызывает глубокие изменения, в особенности у растений, изменяя форму и размеры стебля и листьев, а также обусловливая анатомические изменения органов. На рисунке показано влияние света на наружную морфологию и анатомические признаки дикого латука (Lactuca scariola). При недостаточном освещении изменяется форма стебля, его диаметр становится меньше, появляется полегаемость, листьев мало, они свисают, форма их изменена, листовые пластинки тонкие редуцирована палисадная ткань) и т. п.
Констатировано также, что у наземных моллюсков в условиях сухости и более интенсивного воздействия солнечного света наблюдается увеличение относительного веса раковин. Число «правил» будет расти с расширением исследований.
Влияние химизма питательных веществ и химизма среды также имеет мощное формообразующее значение. Для растений основное значение имеет минеральное питание. Изменения в составе последнего вызывают у них глубокие преобразования формы. Для развития, например, высших растений необходимо присутствие следующих зольных элементов: Ca, Mg, S, Р и Fe. Отсутствие одного из них изменяет формы развития.
Формообразующее воздействие химизма питания на животный организм также очень велико. Неправильно кормленные животные не достигают полного развития, и типичные признаки вида или породы остаются не выраженными. Вообще изменения в химизме среды и ее физических условиях вызывают глубокие превращения формы. Классическим примером остаются результаты экспериментов Шманкевича (1875) и Гаевской (1916), показавшие влияние концентрации солей на формообразование у рачка Artemia. Гаевская показала, что под влиянием убывающей концентрации солей у A. salina происходят изменения в строении брюшка, приводящие к образованию модификантов, сходных по своей внешней морфологической организации с представителями другого рода рачков Branchipus.
Организмы изменяются также под косвенным и прямым воздействием других организмов. Прежде всего это влияние сказывается в процессе конкуренции за источники питания. Сосна, выросшая на свободе, приобретает широкую крону, напоминающую крону дуба, тогда как дуб, выросший в густом лесу, приобретает мачтовый ствол.
Кроме подобных случаев прямого взаимного формообразующего влияния организмов друг на друга, нужно иметь в виду их взаимное косвенное воздействие. Так, например, развитие организмов, в особенности водных, в значительной степени зависит от активной реакции водной среды (концентрации водородных ионов, pH). Большинству водных организмов свойственны известные лимиты приспособленности к определенным пределам pH, а также к известному оптимуму последней, наиболее благоприятствующему развитию. С другой стороны, pH данного водоема в сильнейшей степени зависит от жизнедеятельности находящихся в нем организмов. Так, выделяя СО2, животные вызывают окисление воды, меняя значение pH (в нейтральной воде pH = 7, в кислой pH <7, в щелочной pH >7). стало быть, воздействуя на живущие в воде организмы. Аналогичные формы отношений наблюдаются между растениями, корневая система которых воздействует на минеральный состав почвы (например, бобовые обогащают ее азотом), а следовательно, и на другие растения, развивающиеся в ней.
Состав питательных веществ оказывает формообразующее влияние и на внутренние органы растительных и животных организмов. Быть может, особенно убедительно, преобразующее влияние состава питательных веществ на внутренние органы животных. Одним из классических примеров этого рода является зависимость длины кишечника от растительной и животной пищи. Опыты над головастиками показали, что кормление их животной пищей вызывает уменьшение длины кишечника, воздействуя и на его общую форму.
Перечисленные выше примеры показывают, что внешние факторы вызывают многообразные изменения в организмах.
Характер модификаций. При изучении модификаций была вскрыта весьма характерная черта. Модификации всегда строго закономерны. Модификационные реакции всегда специфичны. Модификационная способность любой формы носит особенный характер. Один и тот же фактор вызывает у разных форм различные модификации, в соответствии с различиями их генотипов (их норм реакции).
Так, повышение температуры вызывает у прыткой ящерицы (Lacerta agilis) посветление кожных покровов (Бидерман, 1892), у стенной ящерицы L. muralis — потемнение (Каммерер, 1906). У антилопы-гну (Африка), по данным М. Завадовского, в условиях асканийской (заповедник Чапли) зимы развивается зимний волосяной покров; у оленебыка (Африка), в тех же условиях, сохраняется летний наряд. Реакция на один и тот же фактор оказывается различной, в зависимости от различия наследственных особенностей данных форм.
Другой особенностью модификационной изменчивости является тот факт, что модификационные изменения одного и того же организма оказываются различными на разных стадиях развития и при разных физиологических состояниях.
Сказанное можно показать на следующем примере. Старые исследования Вейсмана (1895) показали, что у бабочки Araschnia levana наблюдаются две формы: levana и prorsa, различающиеся по рисунку крыла. Первая форма — levana — отрождается из перезимовавших куколок, вторая — prorsa — из летних куколок. В экспериментальных условиях была подтверждена зависимость возникновения этих форм от температурных факторов. Из куколок летней формы при содержании их на холоду вылупляется весенняя форма — levana. Из куколок весенней формы (levana) в тепле вылупляются летние формы prorsa. При дальнейших исследованиях эти зависимости оказались много сложнее. Было показано, что у отдельных индивидуумов наблюдаются различия в развитии куколки.
Одни куколки развиваются непрерывно, тогда как у других наблюдается латентный (скрытый) период, когда видимого развития нет. Если развитие куколки начинается тотчас после окукливания, то при 15—30° из куколки вылупляется летняя форма prorsa. Если развитию куколки предшествует латентный период, то возникает форма levana. Если латентный период затягивается и куколка перезимовывает, то возникает крайняя форма levana. Если латентность длится несколько дней, то развивается форма средняя между levana и prorsa.
С другой стороны, если развитие куколки идет без латентного периода, но при низких температурах (1—10°), то возникает levana. Однако, если низкая температура действует только в течение известного отрезка некоторого чувствительного периода развития куколки, то возникают различные промежуточные формы (Sufflert, 1924). Таким образом, форма модификации в данном случае определяется продолжительностью действия температуры и состоянием самого организма (наличие или отсутствие латентного периода развития). Изложенный пример показывает и третью особенность модификаций — их ненаследственность. Поколение одной и той же формы в разных условиях дает разные модификации.
В. Индивидуальные модификации. Дарвин указывал, что в разных точках семенной коробочки условия различны и, что, следовательно, каждое отдельное семя развивается в индивидуальных условиях. Естественно, что каждое семя окажется индивидуальным модификантом. Все особи, иными словами, обладают индивидуальными модификационными чертами. Этого рода ненаследственную изменчивость мы назовем модификационной (индивидуализацией потомства, или индивидуальными модификациями.
Ненаследственность этих последних в «чистых линиях» самоопылителей попытался показать Иогансен.
Под «чистой линией» понимается серия поколений, полученных от одного конкретного самоопыляющегося растения. Такая «чистая линия» генотипически относительно однородна.
Исследуя «чистые линии» фасоли Phaseolus multifloris, Иогансен показал, что, несмотря на наследственную однородность материала одной «чистой линии», семена потомства ее различны по величине, весу и другим признакам и что эти различия являются последствием индивидуальной диференцировки в условиях развития. Поэтому семена (и другие части растения) модифицируют, и каждое из них отличается от любого другого по весу, размеру и прочим признакам.
Иогансен использовал семена конкретной «чистой линии», чтобы показать ненаследственность модификаций. Он высевал крупные, средние и мелкие семена той же «чистой линии» и констатировал, что размеры семени, в указанных выше условиях, не влияют на размеры семян потомства. Например, от крупных семян получались растения, дававшие и крупные и средние, и мелкие семена. Те же результаты, и притом в пределах той же амплитуды индивидуальных колебаний, были получены при посеве средних и мелких семян. Таким образом, оказалось возможным трактовать индивидуальные модификации одной «чистой линии», как ненаследственные. Иогансен установил также другую черту модификационной индивидуализации, а именно, что она, в пределах каждой «чистой линии», строго закономерна и в частности лимитирована известными, характерными для нее, пределами. Различные «чистые линии» фасоли по Иогансену обладают различными пределами модификационной индивидуализации.
Так как каждая «чистая линия» соответствует конкретному генотипу, то эти данные показывают, что пределы модификационной индивидуализации каждого конкретного генотипа специфичны. Следовательно, процесс модификационной индивидуализации особей одного генотипа служит характеристикой реагирования его на данные конкретные условия, протекая закономерно, вследствие чего явления изменчивости поддаются статистической обработке, излагаемой в курсах генетики.
Процесс модификационной индивидуализации, несомненно, очень важен. Он объясняет непосредственные причины индивидуального многообразия особей, возникающего под влиянием внешних факторов — света, температуры, влажности, питания, химизма почвы, химизма воды и т. п. Под влиянием работ Иогансена в дальнейшем главное внимание генетической мысли было обращено на изучение индивидуальных модификаций. Практически вся проблема ненаследственной изменчивости была сведена к индивидуальным модификациям в «чистых линиях».
F. Однотипные групповые модификации. В пределах системы дарвинизма такая постановка проблемы ненаследственных изменений не удовлетворяет: Нетрудно убедиться, что ненаследственную изменчивость нельзя свести к индивидуальным модификациям. Следует учесть, что особи любого вида связаны видовой общностью, т. е. имеют общее, монофилетическое происхождение. Поэтому, как уже было указано, каждая особь есть единство отдельного и общего. Каждый отдельный генотип есть также единство отдельного и общего. Следовательно, и каждая норма реакции должна быть единством отдельного и общего. Отсюда видно, что всякая модификационная изменчивость должна быть единством отдельной (индивидуальной) и общей (групповой, видовой) изменчивости.
Поясним эту мысль соответствующими примерами. Кожа человека под влиянием солнца загорает. Кожа травяной лягушки под влиянием пониженной температуры темнеет. Мех лисицы, воспитывавшейся в условиях прохладного лета, зимой становится более пушистым и полноволосым. У мышей, воспитанных в тепле, уши длиннее, чем у форм, развивавшихся на холоде, и т. д.
Во всех этих случаях говорится о некоторых модификационных изменениях, носящих однотипно-общий, групповой характер. Но вместе с тем, на фоне этой общей модификации, разыгрывается процесс модификационной индивидуализации, которая имеет то же направление (например, при низком значении температуры все травяные лягушки темнеют, при сильной инсоляции все люди загорают и т. д.), но различное, индивидуализированное выражение (например, все люди загорают в разной степени и в различной форме).
Таким образом, мы будем различать индивидуальные модификации и однотипные с ними массовые, или групповые, модификации.
Установление понятия однотипных групповых (видовых) модификаций с точки зрения системы дарвинизма очень важно. Наличие их показывает, что определенная форма модификационной изменчивости исторически обусловлена, являясь определенным видовым свойством. Во-вторых, их наличие показывает, что наследственное основание каждой особи данного вида также исторически обусловлено и что, следовательно, генотип каждой особи есть единство общего, видового генотипа и индивидуального, отдельного.
Корреляции
Обратимся теперь к явлениям корреляционной изменчивости. Под корреляциями следует понимать вторичные изменения, возникающие в ходе онтогенетического развития под влиянием конкретного первичного изменения. Фенотипически корреляции выражаются в соотносительном изменении функций и структуры органа или ее части, в зависимости от изменений в функции и структуре другого органа или его части. В основе корреляции являются, следовательно, соотносительными функциональными изменениями органов или их частей.
Учение о корреляциях было введено Дарвином в систему дарвинизма отчасти в связи со следующими отношениями. Мы уже знаем, что согласно теории Дарвина эволюция видов идет через процесс их приспособления к меняющимся условиям среды и что расхождение (дивергенция) видов идет по руслу их приспособительной диференцировки.
Казалось бы, в практике исследования виды должны четко различаться по приспособительным признакам. Однако, в действительности этого сплошь и рядом нет. Напротив, в очень большом числе случаев виды значительно явственнее различаются по признакам, приспособительное значение которых не ясно или вообще не может рассматриваться, как таковое.
В любом определителе можно найти десятки примеров, когда именно несущественные в адаптивном отношении признаки имеют наибольшее практическое значение при определении видов.
Систематик не ставит и не может ставить перед собою задачу руководиться непременно приспособительными признаками для различения видов. Он выбирает наиболее четко различаемые признаки, независимо от того, приспособительны они или нет.
Возникает кажущееся противоречие. С одной стороны, расхождение видов достигается путем возникновения приспособительных различий, а с другой, в практике различения (определения) видов сплошь и рядом руководящую роль играют признаки, не имеющие приспособительного значения. Дарвин прямо указывает на то, что именно неприспособительные признаки часто (но не всегда) имеют наибольшее значение в распознавании видов. Разумеется, эти отношения могут быть связаны с нашим незнанием приспособительного значения признаков. Однако факт остается фактом.
Каким же образом возникают эти, более или менее явно неприспособительные признаки различий между видами? Они, казалось бы, не могли быть накоплены отбором, так как отбор накапливает полезные, приспособительные признаки.
Дарвин для объяснения этого кажущегося противоречия привлекает понятие о корреляциях. Он указывает, что ценность несущественных (в смысле их приспособительного значения) признаков для систематики зависит преимущественно от их корреляций с другими, мало заметными, практически недостаточными для определения, но приспособительными признаками. Несущественные в адаптивном отношении признаки возникают, следовательно, не под прямым влиянием отбора, а косвенно, т. е. в силу корреляционной зависимости от других, фенотипически нечетких, но приспособительных признаков. Следовательно, нужно различать ведущие приспособительные изменения и изменения коррелятивные, зависимые. Если возникло приспособительное изменение, то в силу закона корреляции оно влечет за собою возникновение зависимых, коррелятивных признаков. Именно этими зависимыми признаками часто пользуются систематики для более четкого различения видов.
Дарвин поясняет сказанное некоторыми примерами.
Ссылаясь на данные Уаймена, он указывает, что в Виргинии свиньи едят корни одного растения (Lachnantes), причем у белых свиней под влиянием этого растения отпадают копыта, тогда как у черных свиней этого не наблюдается. Поэтому искусственный отбор свиней здесь производится по окраске. Черная окраска коррелятивно связана с адаптивным для данных условий признаком, стойкостью против ядовитых свойств Lachnantes, хотя сама по себе — в условиях искусственного отбора — есть несущественный признак. Селекционер постоянно сталкивается с подобными явлениями. Так, Горленко (1938) указывает, что пшеницы сортов Alborubrum, Milturum, Ferrugineum с красными колосьями в наибольшей степени поражаются черным бактериозом (Bacterium translucens var. indulosum), тогда как белоколосные сорта Velutinum, Hostianum, Nigroaristatum, Barbarossa, Albidum — стойки против этого заболевания. Таким образом, окраска колосьев коррелятивно связана с указанным свойством, хотя сама по себе в условиях селекции не имеет хозяйственного значения.
Это дает право Дарвину подчеркнуть, что вопрос о коррелятивной изменчивости весьма важен, так как, если какие-либо органы изменяются в приспособительном направлении, то вместе с ними изменяются и другие, «без всякой видимой пользы от изменения». Дарвин подчеркивал, что «многие изменения не приносят прямой пользы, но произошли вследствие соотношения с другими более полезными изменениями».
Следовательно, явления корреляции объясняют возникновение и сохранение неадаптивных особенностей в потомстве. Другой важной стороной являлась для Дарвина проблема целостности организма. Изменение в одной части связано с изменениями во всех или многих других частях организма. «Все части организма, — писал Дарвин, — находятся между собою в более или менее тесном соотношении или связи».
Постановка проблемы корреляций в работах Дарвина, а также богатый материал, который был им собран, позволяют определить элементы классификации корреляций между частями организма. Дарвин явственно различал два типа соотношений между частями целостного организма.
Одна группа этих соотношений выражается в существовании особенностей, «которые в обширных группах животных всегда сопровождают друг друга».
Например, все типичные млекопитающие имеют волосяной покров, млечные железы, диафрагму, левую дугу, аорты и т. п. В данном случае речь идет лишь о сосуществовании признаков, по отношению к которым, — пишет Дарвин, — «нам не известно, были ли связаны между собою первичные или начальные изменения этих частей». Описываемые соотношения указывают лишь на то, что «все части тела превосходно координированы для своеобразного образа жизни каждого животного».
Этого рода координации — фактическое сосуществование частей без наличия видимых зависимых связей между ними — Дарвин не рассматривает, как корреляции. Он четко отличает от них «настоящие» коррелятивные изменения, когда возникновение одной части зависит от возникновения другой в ходе индивидуального развития особи. Дарвин собрал большое количество примеров корреляций. Так, изменения в характере роста частей тела моллюска, неравномерный рост правой и левой сторон определяют расположение нервных тяжей и ганглиев у моллюсков и, в частности, развитие их асимметрии; изменения в органах, возникающих на осевом стебле растения, влияют на его форму и т. п.
Отбор голубей-дутышей на увеличение размеров тела привел к увеличению числа позвонков, тогда как ребра стали шире; обратные соотношения возникли у маленьких турманов. У трубастых голубей с их широким хвостом, состоящим из большого числа перьев, заметно увеличены хвостовые позвонки. У гончих голубей длинный язык связан с удлинением клюва и т. п.
Цвет кожи и цвет волос изменяются обыкновенно вместе: «так, уже Виргилий советует пастуху смотреть, чтобы рот и язык у баранов не были черны, иначе ягнята не будут совершенно белыми». Множественность рогов у овец коррелирована с грубой и длинной шерстью; у безрогих коз сравнительно короткое руно; у бесшёрстных египетских собак и безволосого крысодава недостает зубов. Белые кошки с голубыми глазами обычно глухи; пока глаза котят закрыты, они голубого цвета и вместе с тем котята еще не слышат и т. д.
Те же явления у растений. Изменения в листьях сопровождаются изменениями в цветах и плодах; опытные садоводы по листьям сеянцев судят о достоинстве плодов; у змеевидной дыни, плоды которой имеют извилистую форму, около 1 м длиной, стебель, ножка женского цветка и средняя доля листа также удлинены; ярко-красные пеларгонии, имеющие несовершенные листья, имеют и несовершенные цветы и т. д.
Классификация корреляции
Материал, собранный Дарвином, показал многообразие коррелятивных связей и значительный теоретический и практический интерес явления. В последарвиновскую эпоху проблема корреляций разрабатывалась многими авторами второй половины XIX и XX веков.
Не входя в рассмотрение истории классификации корреляций, отметим лишь, что в процессе их изучения многочисленными исследователями была предложена весьма различная терминология. При этом многие из этих исследователей отошли от исторического аспекта на явления корреляций. Мы, прежде всего, вслед за Дарвином, будем строго различать координации и корреляции.
Координациями, соответственно воззрениям Дарвина, следует называть явление сосуществования определенных морфо-физиологических особенностей строения, которые всегда сопровождают друг друга в монофилетических группах видов и скомбинированных в процессе исторического формирования данной группы, причем между координированными частями могут отсутствовать прямые функциональные связи и зависимости.
Такова, например, система признаков или общий «план композиции» типов животного и растительного царства, их классов, отрядов, семейств, родов и т. п. Например, характерные признаки хордовых Chordata — хорда, нервная трубка, жаберные щели в глотке и брюшное положение сердца — образуют координированную систему признаков, постоянно сосуществующих у всех видов хордовых, при всем многообразии их отношений, от низших хордовых до млекопитающих, и притом — от кембрия до геологической современности. У любой особи любого класса, отряда, семейства, рода и вида, в любых естественных условиях, на самых различных отрезках геологического времени указанные выше признаки постоянно сосуществовали (на тех или иных стадиях развития), несмотря на коренные преобразования в других системах органов. Точно так же комбинация признаков подкласса плацентарных млекопитающих — млечные железы, волосяной покров, диафрагма, левая дуга аорты, безъядерные эритроциты, плацентарность и т. п. — существуют у всех особей всех видов, родов, семейств и отрядов этого подкласса, в любых естественных условиях, на любом отрезке геологического времени — от триаса до геологической современности.
Это постоянство сосуществования (известной системы) признаков и означает их координированность. В соответствии с воззрениями Дарвина, крупнейший русский морфолог А. Н. Северцов указал, что «критерием координации мы принимаем признак постоянного сосуществования».
Координации являются результативным итогом накопляющего процесса отбора. Следовательно, координации составляют особую категорию исторических явлений, отличных от любых форм изменчивости. Ввиду этого, проблему координаций мы пока оставим в стороне и обратимся к рассмотрению корреляций, как особой формы изменчивости, т. е. как к одному из источников эволюционного процесса.
Принципы классификации корреляций. Корреляции тесно связаны с онтогенезом и должны рассматриваться, прежде всего, в связи с ним, как всякая форма изменчивости. Вопрос же о роли корреляций в филогенезе рассматривается ниже. Мы примем здесь классификацию И. И. Шмальгаузена (1938).
Поскольку корреляции играют большую роль в ходе онтогенеза, Шмальгаузен придает наибольшее значение классификации корреляций по фазам онтогенеза. С точки зрения системы дарвинизма этот принцип классификации следует считать наиболее правильным. Онтогенез может быть разбит на несколько фаз. В основе онтогенеза организма лежит его генотип. Последний сам по себе не есть арифметическая сумма наследственных факторов. Напротив, последние связаны в нем взаимозависимостями, т. е. скоррелированы и образуют целостную систему наследственных факторов — геном. Каждый генотип есть скоррелированная целостность. Отсюда и возникает представление о геномных корреляциях.
В конкретных условиях среды генотип, развиваясь как целостность, как геном, реализуется в определенном индивидуальном фенотипе.
К геномным корреляциям относятся, прежде всего, некоторые примеры Дарвина. Таковы явления скоррелированности черного цвета шерсти свиней и их стойкости против ядовитых свойств Lachnantes; корреляции между голубыми глазами и глухотой у кошек; между белым цветом шерсти у собак и их немотой, между безрогостью коз и их короткошерстностью; между бесшерстностью и немотой у парагвайских собак. К геномпым корреляциям относится также связь между комолостью баранов прекосов и крипторхизмом (Глембоцкий и Моисеев, 1935); между бесшерстностью и пониженной жизнеспособностью у мышей и т. д.
К этой же группе корреляций нужно отнести соответственные явления у растений. Такова уже указывавшаяся выше корреляция между стойкостью к черному бактериозу и цветом колоса у некоторых пшениц; между зеленым цветом зерен ржи и рядом других признаков — коротким и плотным стеблем, большим числом последних, ранним цветением и скороспелостью и т. п. Непосредственные функциональные зависимости здесь отсутствуют, и связанность перечисленных корреляционных цепей определяется геномными корреляциями.
Морфогенетические корреляции приурочены преимущественно к эмбриональной фазе онтогенеза. Именно на примерах этих корреляций четко выявляется природа корреляционных зависимостей.
С первых стадий развития (дробления) яйца и последующего органогенеза морфогенетические или формообразовательные корреляции играют ведущую роль в эмбриогенезе.
Значение корреляций в формообразовании было доказано большим числом в высшей степени изящных экспериментов, некоторые из которых кратко описываются нами в качестве примеров морфогенетических корреляций.
Если вырезать верхнюю губу бластопора из гаструлы полосатого тритона Triton taeniatus и пересадить ее в эктодерму гаструлы гребенчатого тритона Triton cristatus, например, в области брюха, то в месте пересадки (трансплантация) развивается комплекс спинных осевых органов — нервная трубка и хорда. В результате у зародыша Т. cristatus формируется два комплекса спинных органов — нормальный на спине и другой — на брюхе (Шпеман и Мангольд, 1924). Брюшная сторона была выбрана потому, что нормально на ней перечисленные спинные органы не развиваются. Ясно, что они образуются под формативным влиянием ткани бластопора.
Второй пример. После образования глазной чаши, как известно, развивается хрусталик. Шпеманом (1902), Льюисом (1913), Драгомировым (1929) и другими авторами было выяснено, что при удалении глазной чаши у эмбриона травяной лягушки не образуется и хрусталика. Опыт можно поставить иначе. Если трансплантировать глазную чашу (бокал) в эктодерму, где глаз нормально не развивается, то эта «чужеродная» эктодерма образует хрусталик. Наконец, эксперимент может быть изменен следующим образом. Эктодерма, противолежащая глазной чаше, удаляется, а на ее место имплантируется другая эктодерма. Тогда из материала последней образуется хрусталик (Филатов, 1924). Таким образом, становится ясным, что глазная чаша оказывает формативное действие («организующее» влияние) на образование хрусталика. Однако были выяснены и обратные отношения. Когда хрусталик образовался, то он, в свою очередь, воздействует и на глазную чашу. В присутствии хрусталика она больше, в его отсутствии меньше. Формативное действие чаши, однако, имеет главнейшее значение. Было, например, показано (Попов, 1937), что под индуцирующим влиянием глазной чаши линза образуется из закладки нервной системы или мышц, т. е. в среде тканей, которым линзообразование совершенно не свойственно.
Аналогичные явления наблюдались и по отношению к развитию слуховых пузырьков. Если пересадить на брюхо эмбриона тритона кусочек бластопора, то образуется нервная (медуллярная) пластинка, и вслед затем, по ее бокам, как правило, начинается развитие слуховых пузырьков. Следовательно, нервная пластинка индуцирует их образование. Далее, Филатовым было выяснено, что если пересадить слуховой пузырек жабы в область тела, где нормально ухо не развивается, вокруг имплантированного слухового пузырька начинается образование слуховой хрящевой капсулы. Таким образом, слуховой пузырек оказывает формативное действие на возникновение слуховой капсулы.
Эти данные приводят к следующему выводу: за конкретный процесс формообразования ответственны какие-то формообразующие вещества, которые развиваются в закладывающихся органах. Действительно, формативное действие оказывают особые вещества химической природы. Если убить ткань губы бластопора нагреванием, спиртом и т. п., то имплантация этой мертвой ткани оказывает прежнее формативное действие.
Изложенные выше опыты целым рядом исследователей были распространены на самые различные части эмбриона, причем во всех случаях были ясно показаны корреляционные зависимости между органами. Оказалось, что можно говорить о развитии «последовательных звеньев корреляционной цепи». (Шмальгаузен, 1938). Так, зачаток верхней губы бластопора индуцирует образование хорды и нервной трубки; развитие головного мозга стимулирует развитие глазной чаши; последняя вызывает образование линзы; линза индуцирует превращение противолежащей эктодермы в прозрачную роговицу; с другой стороны, закладка мозга влечет за собою развитие слухового пузырька, зачаток последнего оказывает формативное действие на слуховую капсулу и т. п. Этого рода морфогенетические корреляции могут быть названы ступенчатыми (Шмидт, 1938).
Во всех разобранных случаях условием развития всякой последующей части является относительно тесный контакт ее с предшествующей частью, оказывающей на нее формативное действие. Можно, следовательно, говорить о контактных морфогенетических корреляциях, которые играют большую роль в формообразовании органов. Их форма, положение, размеры, дефинитивная моделировка определяются этим контактным воздействием предшествующей части. «Формообразовательная реакция» (Филатов) индуцируемой части определяется, таким образом, «формативным действием» индуктора. Например, глазная чаша, имея значение индуктора, оказывает формативное действие на индуцируемую эктодерму, формативная реакция которой выражается в образовании хрусталика. Подобные контактные коррелятивные связи охватывают многие органы. Контакт между частями организма действует и механически, и биохимически.
В других случаях непосредственного контакта между частями нет, однако формативное действие все же имеется. В этих случаях вопрос идет о связях и корреляциях, которые мы, для краткости, назовем инконтактными. Примером их может служить гормональное формативное воздействие эндокринных желез на органы, воспринимающие эти воздействия. Гормоны (половых желез, щитовидной железы, гипофизы и т. п.) влияют на соответственные органы или части тела путем транспортировки гормональных веществ через кровяное русло. Примером может служить воздействие половых гормонов на сложный комплекс вторичнополовых признаков самки и самца.
Морфогенетические коррелятивные изменения происходят вследствие возникновения первичных изменений, которые влекут за собою соответственные зависимые вторичные изменения. Это явление было доказано экспериментально. Если у зародыша тритона или лягушки вырезать участок нервной трубки с подлежащей крышей первичной кишки и затем вставить в рану этот же кусочек. Но повернув его на 180°, то впоследствии изменяется нормальная топография органов: органы, нормально развивающиеся слева, возникают справа, и наоборот. Возникает обратное расположение органов (situs iniversum). Следовательно, первичное изменение (поворот крыши первичной кишки на 180°) вызвал зависимое вторичное изменение.
Эргонтические корреляции, по преимуществу, относятся к постэмбриональному периоду онтогенеза, но особенно характерны для ювенального периода. Их значение состоит в окончательной моделировке индуцируемых частей. Эргон по-гречески означает: работа. Эргонтические, или рабочие, корреляции возникают обычно в результате контакта между соответственными частями организма. Особенно отчетливо выявляются эргонтические корреляции во взаимоотношениях между работающей мышцей и подлежащей костной опорой. Известно, что чем сильнее развита мышца, тем резче развиваются гребни на участках кости, к которым она прикрепляется. Поэтому по степени развития костных гребней можно судить о степени развития мышцы, служащей эргонтическим индуктором, формативно воздействующим на кость.
Эти отношения особенно четко проявляются в моделировке черепа млекопитающих. Если проследить формирование черепа на протяжении ювенальной и взрослой фаз, то легко обнаруживается воздействие работающей мускулатуры на пластику черепа. По мере развития теменной мышцы по бокам черепа образуются теменные линии (linea temporalis). По мере роста теменной мышцы по направлению к сагиттальному шву черепа, теменные линии, в результате зависимой перестройки костного вещества, передвигаются к сагиттальному шву и, встречаясь здесь, как две волны, образуют высокий сагиттальный гребень.
Твердый костный череп оказывается необычайно пластичным. Не череп определяет форму мозга, а мозг накладывает свою печать на конфигурацию черепа. Не череп определяет форму мышц, а они воздействуют на него. Эти эргонтические корреляции, в данных примерах — зависимость формы костного вещества от функции мышц — были доказаны экспериментально. Было, например, доказано, что симметрия черепа — следствие симметричной функции жевательной мускулатуры. В правой нижней челюсти зубной ряд, напротив, стерт больше, чем в левой нижней челюсти. Эти данные показывают, что животное работало нижней челюстью неравномерно и, возможно, в несколько скошенном напра^ влении, изнашивая зубной ряд верхней левой челюсти и нижней правой челюсти. Эти отношения связаны с асимметричной функцией жевательных мышц. С правой (дефектной) стороны усилилась деятельность теменной мышцы, которая развилась здесь сильнее. В зависимости от этого правая теменная линия придвинулась вплотную к сагиттальному шву.
Слева оказалась усиленной деятельность другой жевательной мышцы, m. masseter, прикрепляющейся одним своим концом к скуловой дуге, а другим к основной части восходящей рамы нижней челюсти. Интенсивная работа этой мышцы вызвала коррелятивное изменение в структуре восходящей ветви нижней челюсти, а именно более резкое углубление места ее прикрепления. В то же время интересное изменение произошло в строении скуловой дуги, на нижнем крае которой образовался отросток, несвойственный черепам хищников. Таким образом, мы получаем следующий общий ход изменений:
1) правая верхняя челюсть (вследствие какого-то поражения) росла по всем направлениям медленнее левой; 2) вследствие этого череп искривился; 3) изменился способ жевания; 4) возникла асимметрия функции жевательных мышц; 5) изменилось строение места прикрепления m. masseter в левой нижней челюсти; 6) образовался несвойственный хищникам отросток скуловой дуги.
Мы видим, следовательно, что под влиянием первичного изменения (1) возникли зависимые вторичные изменения (2—3), в данном примере, — эргонтические корреляционные изменения в строении нижней челюсти и скуловой дуги. Мы можем теперь совершенно четко установить, что эти изменения — прямое следствие коррелятивных отношений, определенных зависимостей и условий, возникших в системе развивающегося черепа. Аналогичный вывод касается всех типов коррелятивных связей и, следовательно, всех признаков органической формы. Во времени между генотипом и конечными фенотипическими признаками организма лежит область формообразовательных процессов онтогенеза, которые связаны сложными корреляционными цепями. Генотип определяет только многообразные возможности онтогенетического развития, только норму реакции конкретного организма. Фенотипические признаки, как таковые, формируются под влиянием условий развития (модификации) и зависимых коррелятивных изменений.
Модификации, мутации и корреляции создают, таким образом, огромное многообразие индивидуумов, причем потенциальные возможности этого многообразия далеко не исчерпываются.
Причины изменчивости
В возникновении изменений ведущую роль играют внешние, по отношению к данному организму, причины. По отношению к модификациям это положение не требует особых объяснений. Мы видели, что модификации являются реакциями на внешние по отношению к организму воздействия, определяемые генотипической нормой реакции. В отношении корреляций вопрос, как будто, осложняется. Корреляционные изменения возникают, как мы видели, под влиянием внутренних, в самом организме развивающихся соотношений между органами и их частями. Однако и в отношении корреляций ясно, что зависимые изменения, протекающие в каком-либо органе или его части (например, в хрусталике глаза), являются не чем иным, как реакцией на внешние, по отношению к этому органу или его части, воздействия. Всякое первичное изменение в одном органе индуцирует изменения в другом. В пределах одного и того же генотипа вторичного корреляционного изменения не произойдет, если не было первичного. Следовательно, корреляции возникают по типу эктогенетического процесса и могут рассматриваться как особый тип модификационной изменчивости, определяемой внутренней средой системы самого организма. Только в случае изменения самого генотипа, а следовательно, и нормы реакции, другими словами, — в случае мутации, — характер модификаций и формы корреляций изменяются. Следовательно, в основном, проблема причин изменчивости упирается в вопрос о причинах наследственных (мутационных) изменений.
Де-Фриз, которому принадлежит самый термин мутация, исходил из неверного положения, что наследственные изменения возникают вне зависимости от внешних факторов. Он предполагал, что всякой мутации предшествует некий автономный «премутационный период». Эта точка зрения должна быть названа автогенетической. Идея автогенеза получила широкое распространение в генетике и вызвала оживленную борьбу между автогенетиками и эктогенетиками, считающими, что причину наследственных изменений нужно искать во внешних факторах.
Обращаясь к вопросу о влиянии внешних факторов на возникновение наследственных изменений, необходимо, Прежде всего, уяснить себе тот факт, что эктогенетическая точка зрения не должна смешиваться с механицизмом. Механистическое воззрение сводит проблему возникновения наследственных изменений под влиянием внешних факторов только к этим последним, не учитывая специфики развития организма. В действительности, неверно утверждать, что, наследственное основание организма изменяется под влиянием внешних факторов пассивно. Дарвин неоднократно подчеркивал, что в возникновении изменений играют роль как внешние факторы, так и природа самого организма. Морфофизиологические свойства организма и его биохимическая структура играют решающую роль в том, каковы будут наследственные изменения его под влиянием внешних факторов. Проникнув в организм, внешний фактор перестает быть внешним. Он действует в физиологической системе организма как новый внутренний фактор. «Внешнее» становится «внутренним».
Поэтому возникновение наследственных изменений никогда не бывает анархичным; — оно всегда закономерно.
Для доказательства рассмотрим следующие данные. Впервые экспериментально мутации были получены Мёллером (1927), применившим для этой цели воздействие рентгена. Им и последующими авторами было доказано, что под влиянием рентгена у плодовой мушки могут быть получены мутационные изменения антенн, глаз, щетинок на теле, крыльев, размеров тела, окраски, степени плодовитости, продолжительности жизни и т. п. Следовательно, один и тот же фактор вызвал различные наследственные изменения. Направления наследственных изменений конкретного организма определяются не внешними факторами, а самим организмом.
Очень удачен несправедливо забытый термин Дарвина «наследственная индивидуальная неопределенная изменчивость». Внешний фактор не определяет ее направления, она остается единичной, объективно случайной, неопределенной.
Вопрос, разбираемый нами, может быть проверен и с другой стороны. Если один и тот же фактор вызывает у конкретного организма разные изменения, то, с другой стороны, в ряде случаев различные факторы вызывают одни и те же наследственные изменения. Например, у львиного зева Antirhinum majus температура, ультрафиолетовое облучение, химические агенты вызывали сходные мутации — карликовый рост, узколистность и т. п.
Наконец, должно быть подчеркнуто, что разные видовые формы реагируют, в смысле их наследственной изменчивости, различно, обладая, в частности, различной чувствительностью по отношению к одному и тому же фактору. Например, в одинаковых условиях эксперимента один вид плодовой мушки Drosophila melaniogaster мутируется легче, чем другой — Dr. funebris. У разных пшениц, несмотря на одинаковые условия эксперимента, наблюдается та же картина. Совокупность, описанных фактов подтверждает, что наследственная изменчивость идет в разных направлениях закономерно.
Итак, возникает следующее представление о причинах наследственных изменений.
1. Внешние факторы играют роль индукторов, вызывающих наследственные изменения.
2. Наследственная специфичность организма определяет направления изменчивости.
Индукторы наследственной изменчивости
Внешние факторы, вызывающие наследственные изменения (мутагенные факторы), могут быть распределены по двум группам: искусственные и естественные (действующие в природной обстановке). Деление это, разумеется, условно, однако, оно представляет некоторые удобства.
Рассмотрим, прежде всего, влияние рентгена. Выше упоминалось о мутагенном действии его. Облучение рентгеном вызывает усиление мутационного процесса у самых различных организмов. Его мутагенное действие было доказано на дрозофиле, вощинной моли, на осе Habrobracon juglandis, на ряде растений — хлопчатнике, кукурузе, ячмене, овсе, пшенице, ржи, львином зеве, томате, табаке, гиацинте и т. п. Облучение вызывало появление у дрозофил ряда мутаций в различных органах. У Habrobracon было получено 36 мутаций (Уайтинг, 1933) жилкования, формы и величины крыльев, окраски и формы тела, окраски и формы глаз и т. п. Как значительны получаемые радиоактивным воздействием изменения, показывает рисунок, где изображены исходные формы пшеницы albidum 0604 и ее мутации. Мутагенное действие радия испытано на многих растениях. Так, отчетливые мутации формы листьев, окраски и величины растений были получены при воздействии радием на покоящийся зародыш львиного зева. Большое число работ, посвященных проблеме мутагенного значения рентгена и радия (Бебкук и Коллинс, 1929; Hanson и Heys, 1929; Иоллос, 1937; Тимофеев-Рессовский, 1931 и др.), позволяет, однако, утверждать, что естественная радиация во много раз слабее той, которая имеет мутагенное значение. Таким образом, видеть в рентгене и радии причину естественных мутаций едва ли возможно.
Ультрафиолетовые лучи также имеют мутагенное действие. Так, действием ультрафиолетовых лучей удалось получить мутанта инфузории Chilodon uncinatus (Мак Дугалл, 1931), отличающегося от нормальных форм наличием хвостового выроста. Были получены также мутации у дрозофилы (Альтенберг, 1930). Облучение почек львиного зева (Штуббе, 1930) имело результатом появление узколистных и карликовых форм.
Мутагенное влияние химических веществ было очень четко показано Сахаровым (1932), получившим мутации у дрозофилы под воздействием йодистых препаратов на яйца мух. Аналогичные данные получили Замятина и Попова (1934). Гершензон (1940) получил мутации в строении крыльев дрозофилы в результате кормления личинок натриевой солью тимонуклеиновой кислоты. Баур (1930) воздействовал на семена львиного зева хлоралгидратом, этиловым спиртом и другими: веществами, доказав их мутагенное действие.
Температура также оказывает мутагенное действие. Нужно считать, что мутагенное влияние температуры было доказано еще Тоуэром (1906), экспериментировавшим с картофельным жуком Leptinotarsa, воздействуя повышенной температурой на жуков в период вызревания половых продуктов. Тоуэр получил несколько мутантных форм, отличавшихся окраской и рисунком элитр и спинки. При скрещивании мутантов с нормальными формами в первом поколении были получены формы, фенотипически соответствующие нормальным. Однако, во втором поколении наблюдалось расщепление. Так, в одном опыте во втором поколении оказалось 75% нормальных форм (L. decemlineata) и 25% мутантов типа pallida. Таким образом, полученные изменения оказались наследственными и должны рассматриваться как мутации.
Гольдшмидт (1929) исследовал также мутагенное влияние температуры на плодовых мухах. Была использована сублетальная (близкая к смертельной, или летальной) температура 37°, действовавшая в течение 10—12 час. Опыт вызвал большую смертность, но, с другой стороны, был получен ряд мутантных форм. Подобные же опыты были проведены Иоллосом (1931, 1934, 1935), получившим мутации окраски глаз.
Интерес к проблеме мутагенного влияния температуры, как естественного фактора, стимулировал дальнейшие исследования, причем было доказано (Биркина, 1938; Готчевский, 1932, 1934, Зуйтин, 1937. 1938; Керкис, 1939 и другие авторы), что температурный фактор, безусловно, имеет мутагенное значение, хотя частота мутаций под его влиянием ниже, чем, например, под влиянием рентгенизации.
Что касается растений, то наиболее отчетливые результаты были получены Шкварниковым и Навашиным (1933, 1935). Эти авторы показали, прежде всего, что высокие температуры дают огромное повышение частоты мутационного процесса. Авторы экспериментировали с семенами скерды (Crepis tectorum) и пшеницы 0194 Одесской селекционной станции. При этом было изучено влияние различных температур в условиях различной продолжительности их действия. В частности, были констатированы альбиносные растения.
Температурный фактор играет столь большую роль в природе, что эти данные представляют значительный интерес, подчеркивая справедливость представления о естественном возникновении мутаций под влиянием природных факторов.
В этой связи приобретают большой интерес работы, авторы которых стремятся вынести эксперимент в пределы естественной обстановки, экспериментировать в природе. Остановимся здесь на недавних работах Сахарова и Зуйтина. Сахаров (1941) исследовал влияние зимовки в условиях пониженной температуры на плодовых мух Drosophila melanogaster. В частности, он установил, что 40—50-дневная зимовка самок и 50—60-дневная зимовка самцов привели к явственному повышению частоты мутационного процесса, особенно в потомстве тех особей, среди которых в результате тяжелой зимовки, наблюдалась массовая гибель. Сахаров пришел к выводу, что, повидимому, накопление мутаций во время зимовки «является одним из факторов, ведущих к повышению внутривидовой наследственной изменчивости». В следующей главе мы еще вернемся к его данным.
Зуйтин (1940) исследовал влияние на мутационный процесс у плодовых мух замены лабораторных условий развития природными. Зуйтин ожидал, что замена устойчивого лабораторного режима естественным, отличающимся колебаниями условий и снижением общего уровня температуры, сама по себе должна повлиять на частоту мутационного процесса. Для проверки этого предположения лабораторная культура дрозофил была завезена на Кавказ (Сухуми и Орджоникидзе) и выпущена здесь в природную среду. Для предотвращения скрещивания с местными мухами, привезенная культура была изолирована при помощи марли. Мухи находились в условиях колеблющейся влажности и температуры. Эти отношения сказались на развитии их потомства. Зуйтин констатировал повышение частоты мутационного процесса. В другой работе (1941) Зуйтин исследовал влияние контрастов влажности на Dr. melanogaster. Он установил, что резкое снижение влажности в начальный период развития куколок привело к существенному увеличению процента мутаций.
В той же связи интересны результаты исследования, ставившего целью выяснить мутагенное значение пищевого режима. Было, например, показано, что нарушение режима питания львиного зева повышает частоту мутирования. Таким образом, доказано, что естественные факторы (температура, влажность и т. п.) ответственны за возникновение мутационного процесса. Действие внешних факторов на наследственное основание организма имеет, однако, сложный характер. Мутационный процесс возникает и в условиях относительно постоянной среды, под влиянием обменных процессов в клетках. Мутационный процесс есть выражение развития генома. Рассмотрение этого вопроса связано, однако, с проблемой наследственности.