До сих пор мы рассматривали процессы, обусловливающие возникновение, наследование и поддержание различий внутри популяции.
Теперь остановимся на процессах, ведущих к появлению межпопуляционных различий и зарождению внутри вида новых форм, рас, подвидов, т. е. рассмотрим начальные этапы процесса видообразования.
Процесс видообразования, связанный с пространственным разобщением популяций, носит название аллопатрического видообразования, тогда как возникновение новых форм без пространственного разобщения называют симпатрическим видообразованием.
Некоторые исследователи полагают, что аллопатрическое видообразование представляет собой основной путь становления новых видов. Результаты изучения эколого-географических связей между близкими видами в большинстве случаев с несомненностью указывают на их первоначальную пространственную разобщенность.
Рассмотрим некоторые примеры аллопатрического видообразования. Из 1200 видов рыб к востоку и западу от Панамского перешейка, который образовался в геологически недавнее время, только 6% оказываются общими для фауны обоих океанов — Атлантического и Тихого, тогда как многие остальные виды представляют собой викарирующие, т. е. географически разобщенные и давно дивертировавшие формы. Сходная картина наблюдается здесь и в отношении животных других классов — моллюсков, морских ежей, червей, ракообразных и т. д.
Б. К. Шишкиным описаны 3 географические расы (подвиды) лесного купыря (Anthriscus silvestris) — европейская, кавказская и сибирская, каждая из которых обладает своим приспособительным комплексом морфоэкологических особенностей. Другое растение нашей флоры — прострел (Anemone pulsatilla) занимает непрерывный ареал по всей Европе с запада на восток, но при этом для растений на западе характерны сильно рассеченные листья и поникшие цветки, а для растений на востоке — слабо рассеченные листья и стоячие цветки.
Согласно Ч. Дарвину, широта ареала непосредственно способствует процессу видообразования. Географические расы или подвиды в некоторых случаях могут рассматриваться как новые виды в потенции, однако для реализации этих потенций и приобретения подвидами видового ранга требуется их дальнейшая дивергенция, приводящая в конечном счете к нескрещиваемости, репродуктивной изоляции, возникновению различных адаптивных механизмов.
Географические расы всегда оказываются в различающихся условиях, что определяет их большую или меньшую экологическую специализацию. Поэтому они могут одновременно рассматриваться и как экологические расы. Хорошим примером экологической специализации в связи с пространственным разобщением могут служить так называемые «кольцевые» виды. Ареалы серебристой чайки (Larus argentatus) и клуши (L. fuscus) в Западной Европе перекрываются. Оба эти вида четко различаются друг от друга и не скрещиваются. Однако если проследить их географические расы к западу (Гренландия, Лабрадор, Канада, Аляска) и к востоку (Северная Европа, Северо-Западная и Северо-Восточная Сибирь), то выявится, что последние образуют непрерывное циркумполярное кольцо вокруг Ледовитого океана. При этом скрещивание между особями соседних географических рас происходит свободно. Н. В. Тимофеев-Ресовский предполагает, что первоначальное расселение чаек к западу и востоку началось с какого-то промежуточного района, например с Восточной Сибири. Оно сопровождалось усиливающейся специализацией географических рас, так что когда кольцо в Западной Европе замкнулось, родственные формы (L. argentatus и L. fuscus) «не узнали» друг друга, оказались разными видами.
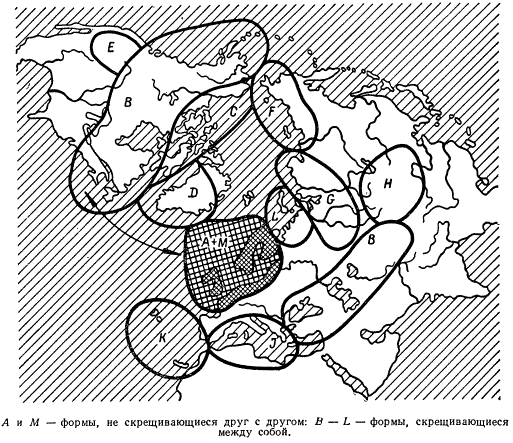
Географическое размещение чаек
Экологическая специализация внутри вида может иметь место и в смежных областях без существенного географического удаления дивергирующих форм, при сохранении возможности обмена генетическим материалом между особями из соседних популяций. Еще Дарвином был описан яркий пример пищевой специализации вьюрков рода Geospiza, обитающих на Галапагосских островах и разошедшихся по размеру и форме клюва. Показательно также в этом отношении обилие эндемичных форм в крупных закрытых водоемах. Так, в оз. Байкал обитает более 300 видов ракообразных, относящихся к семейству Gammaridae, большинство из которых нигде больше не встречается. В оз. Ньяса найдено 174 эндемичных вида рыб семейства Cichlidae, а в оз. Танганайка — около 80 эндемичных видов рыб семейства Chromidae. Возникновение всех этих эндемов обусловлено, по всей вероятности, экологической изоляцией, различиями в направлении действия отбора в локально обособившиеся популяциях.
Экологическая специализация определяет, очевидно, и формирование экотипов у растений, приуроченных к определенным экологическим нишам и характеризующихся наследственно закрепленным комплексом морфо-физиологических особенностей.
К рассматриваемой категории явлений следует отнести также сезонную изоляцию, проявляющуюся в расхождении по срокам размножения. Л. С. Берг выделил 5 форм форели (Salmo ischchan) оз. Севан, существенно различающихся по срокам икрометания и некоторым другим биологическим особенностям. Им же установлены «яровые» и «озимые» расы рыб из семейств лососевых, осетровых, карповых, нерестящихся соответственно весной или осенью. У одних и тех же видов растений есть яровые и озимые формы, а также расы-эфемеры, цветущие еще весной, и позднеспелые, зацветающие лишь под осень. И во всех случаях возникновение внутривидовой дивергенции, связанное с экологической специализацией, способствует подгонке популяций к конкретным условиям существования и обеспечивает наиболее полное использование территориальных, пищевых и прочих ресурсов района обитания.
В основе расхождения по морфофизиологическим особенностям географических или экологических рас лежит дивергенция генетической структуры составляющих их популяций. Если такая дивергенция заходит достаточно далеко, то это приводит к развитию механизмов репродуктивной изоляции, способных в той или иной степени ограничивать возможность скрещивания особей, относящихся к различным расам.
Основные формы репродуктивной изоляции выражаются:
- в изменении характера избирательности спаривания,
- в изменении характера избирательности оплодотворения,
- в понижении фертильности гибридных особей,
- в снижении жизнеспособности гибридов.
Изменение характера избирательности спаривания (поведенческая изоляция) часто предшествует в своем возникновении у животных появлению других форм репродуктивной изоляции. Это связано с тем, что половое поведение особей противоположного пола представляет собой сложный, высокоспецифичный в видовом отношении, тонко скоординированный комплекс наследственно контролируемых (безусловных) реакций. Примером этого могут служить брачные игры у многих видов рыб, птиц и млекопитающих, сопровождающиеся демонстрацией яркого брачного наряда. Весь этот комплекс реакций обеспечивает, с одной стороны, синхронизацию половых циклов самцов и самок, а с другой — предотвращение межвидовых скрещиваний, т. е. обеспечение репродуктивной изоляции.
О наличии избирательности оплодотворения у животных судят на основе опытов, в которых сопоставляется оплодотворяющая способность спермы самцов из разных линий или пород при скрещивании их с самками разного генотипа; у растений наличие избирательности оплодотворения выявляют по конкурентоспособности пыльцевых трубок при опылении смесью пыльцы. Что же касается двух других механизмов репродуктивной изоляции — понижения фертильности и жизнеспособности гибридов, то их возникновение отражает, как правило, результат далеко зашедших генетических различий, когда дивергировавшие формы достигают ранга самостоятельных видов.
Для симпатрического видообразования не обязательно пространственное разобщение популяций. У животных и растений существует целый ряд механизмов, которые могут обеспечить внутривидовую дивергенцию. Уже в наследственно обусловленном фенотипическом полиморфизме популяций, в основе которого лежит разнообразие условий обитания, содержатся предпосылки для симпатрического образования новых форм. Это доказывается существованием в одних и тех же географических районах и местообитаниях близких видов бабочек, тлей и др., каждый из которых является строгим монофагом, т. е. питается на своем определенном растении-хозяине. Среди паразитирующих простейших также известны близкие формы, поражающие разных хозяев.
У растений существенными генетическими факторами симпатрического видообразования могут быть полиплоидия, мутации, определяющие несовместимость геномов и цитоплазмы. У животных такими факторами являются гетерогаметность пола, гены стерильности и даже цитоплазматическая изоляция. Примером последнего может служить следующее явление, установленное Г. Лавеном на комарах (комплекс Culex pipiens), которые широко распространены в Европе, Азии и Северной Америке. При скрещивании двух легко скрещивающихся форм из Южной (Og) и Северной Германии (На) некоторые комбинации гибридов почти не появлялись, так как оплодотворенные яйца развивались только до какой-то эмбриональной стадии, а затем гибли.
Все гибриды последнего скрещивания (0,17%) оказались самками и содержали только материнский геном, т. е. развивались партеногенетически. Сперматозоиды (На) лишь стимулируют развитие. С помощью серии беккроссов [(На X Og) X Og] X Og и далее было проведено практическое замещение всех трех пар хромосом формы На на хромосомы формы Og. Было проведено более 50 поколений указанных беккроссов, результаты неизменно соответствовали скрещиванию На X Og. Отсюда был сделан вывод, что способность вызывать несовместимость при скрещивании og X На обусловлена не ядерными генами, а цитоплазмой. Оказалось, что цитоплазма формы hfa определяет неспособность спермиев этой формы оплодотворять яйца формы Og.
Теперь предположим, что в популяции формы Qg в результате мутации возникла особь, сходная с формой На. Если эта особь оказывается самцом, то она может оплодотворять только самок Og, но при этом никакого потомства не образуется. Если мутации будет нести самка, то, оплодотворенная самцами Og, она даст самок и самцов, но последние будут проявлять свойства На. Возникшая небольшая группа особей типа На в дальнейшем будет увеличиваться как за счет размножения внутри себя, так и за счет оплодотворения самок На самцами Og. При этом за счет скрещивания Og X На численность популяции Og сократится, т. е. произойдет замещение одной генетической структуры на другую (Og на На).
Цитоплазматическая изоляция может служить не только механизмом симпатрического видообразования, но, возможно, и методом борьбы с насекомыми — переносчиками болезней путем запуска в природу определенного типа самцов.
В процессе симпатрического видообразования могут иметь значение онтогенетические механизмы адаптации и индивидуального опыта у значительной группы животных, базирующегося на выработке в течение жизни разнообразных условных рефлексов, которые могут быть переданы затем потомству с помощью механизма сигнальной наследственности. Хорошо известны факты гнездового консерватизма птиц, при котором одни и те же особи каждый раз возвращаются для размножения в одни и те же места. Во всех подобных случаях животные по сигналам лишь выбирают определенные сочетания факторов среды, что может привести к стойкому расчленению и обособлению ранее единой популяции.
Выработка половых условных рефлексов у птиц способна также обеспечивать чуть ли не полную избирательность спаривания самцов с самками, с той или другой окраской оперения. Этим путем может осуществляться направленное формирование репродуктивной изоляции за счет отдельных мутантных признаков.
Все приведенные выше факты свидетельствуют о том, что симпатрическое видообразование есть реальный способ формирования новых видов, который по масштабам своего действия в природе не только не уступает аллопатрическому видообразованию, но, по-видимому, для животных как организмов, обладающих способностью к передвижению и активному приспособлению к среде, является основным. Но каково бы ни было истинное соотношение аллопатрического и симпатрического видообразования, необходимо иметь в виду, что решающее значение во внутривидовой дивергенции и в возникновении изолирующих механизмов остается за действием отбора и генетическими механизмами, ведущими и направляющими процессы микроэволюции.
Итак, генетико-популяционный подход в сочетании с методами цитогенетики, экологии, физиологии и математической генетики позволил вскрыть некоторые существенные моменты процесса микроэволюции.
С очевидностью вытекает, что факторы эволюции — изменчивость, наследственность и отбор нельзя рассматривать как независимые процессы. Они тесно связаны между собой и взаимозависимы. Путем отбора можно создать линии организмов с большей или меньшей наследственной изменчивостью. Могущество естественного и искусственного отбора беспредельно.
Подвергая организмы действию мутагенов (температуры, химических веществ и др.) в ряду поколений и ведя отбор на устойчивость к ним, можно создавать линии, которые будут обладать различной мутабильностью под действием данного мутагена. Между факторами отбора и наследственной изменчивостью, по-видимому, существует прямая зависимость: если резко меняются условия внешней среды, то вид имеет возможность приспособиться к ним либо путем использования имеющегося мутационного резерва, либо за счет увеличения частоты возникновения новых мутаций.
Хотя взаимодействие факторов эволюции кажется очевидным, конкретные механизмы этой связи изучены очень слабо. Эти вопросы подлежат непременному и быстрейшему исследованию, поскольку решение их необходимо не только для познания эволюции, но и для создания новых форм с помощью методов селекции.