В список экологических факторов принято помещать рельеф (орографические факторы), а иногда и возраст (исторические факторы), так как возраст и рельеф оказывают влияние на лес.
Констатируя влияние рельефа на растения, мы тем самым указываем на различия пространственного распределения среды. Рельеф перераспределяет в пространстве экологические факторы — свет, тепло, влагу, питательные вещества почвы. По отношению к растению он является распределителем факторов среды, именно — пространственным распределителем.
Когда исследуют влияние рельефа на рост леса в конкретных случаях, то рельеф мыслят как горы, равнины, речные долины, склоны и т. п., т. е. как пространственно разнообразные комплексы почв, горных пород, климатических факторов, увлажнения и т. д. Когда отмечают, например, что черная ольха приурочена к пониженному рельефу, то этим самым указывают на пониженный рельеф как на фактор, создающий благоприятные условия роста для ольхи (увлажнение, особый режим минерального питания и другие конкретные особенности местообитания).
Равным образом и экологический фактор времени нельзя представлять себе иначе, как процесс изменения растений и среды, происходящий в связи с их ростом и развитием. Появление самосева и подроста, образование чащи-молодняка, дальнейший переход молодняка в жердняк, затем в средневозрастное, приспевающее и спелое насаждение — все это примеры действия фактора времени. Это действие заключается в процессах роста и развития леса, усвоения деревьями их среды — пищи, нарастания древесины, плодоношения и других процессов, вызывающих переход насаждений из одной стадии в другую.
В дальнейшем на примерах борового и других лесных комплексов будет показан характер связи факторов среды между собой, их взаимодействие с растениями и особое значение так называемых ведущих факторов среды.
Для того чтобы лучше разобраться в сложной логике упоминаемых связей, следует обратиться к наиболее простым и наглядным примерам. Проще и доходчивее постигаются они в том случае, когда берут пример зависимости изменений растительности от количественных изменений какого-либо одного фактора среды при неизменном количестве всех остальных. Такого рода пример легче всего воспроизвести в искусственной обстановке, в условиях вегетационных пли полевых опытов, когда один и тот же вид пли сорт растения культивируют на одинаковых субстратах и почвах, дозируя один и тот же фактор (например, влагу или питательные вещества — азот, фосфор, калий и т. п.) в определенных нарастающих или убывающих количествах. В таком случае полученную разницу в росте и развитии растений объясняют влиянием дозируемого (изменяемого) фактора среды.
В условиях леса получить подобную возможность весьма трудно. Приходится искать такие комплексы или экологические ряды, которые в какой-то степени удовлетворяли бы требованиям ряда с одним меняющимся фактором. Совершенно ясно, что таких простых, чистых зависимостей, способных дать точность учета до 1%, в природе не существует. Приходится довольствоваться экологическими комплексами и рядами, на материале которых можно изучать влияние отдельных факторов как ведущих на данном участке территории в данном экологическом ряду. В таком случае задача облегчается и становится разрешимой при помощи обычных геоботанических, почвенных и лесоводственных методов.
Анализ экологических рядов лесных биоценозов иллюстрирует качественно-количественные переходы в системе «организм — среда».
Остановимся на примере профиля через боровой комплекс. Он представляет собой экологический ряд самых бедных песчаных почв и соответствующих им насаждений — сосняков, простых по составу и взаимоотношениям. На рисунке схематически показан характерный для южной подзоны хвойных лесов (в частности, для Полесья) профиль, расположенный вдоль склона песчаной возвышенности.
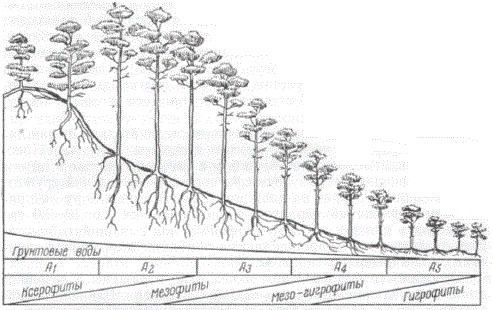
Схема борового экологического ряда сосняков на склоне дюны от сухого бора A1 на ее вершине до сосны по болоту А5 у ее подножия
В типичном профиле такого рода насаждения состоят только из двух наименее требовательных к почве древесных пород — сосны и березы. На вершине бугра — сухой (лишайниковый) бор из чистой сосны III—IV бонитета А1, переходящий далее вниз по склону в свежий бор (зеленомошник-брусничник) I—II бонитета А2. Свежий бор сменяется влажным бором — зеленомошником-черничником III бонитета А3, а этот последний переходит в сырой бор (долгомошник) IV бонитета с черникой, голубикой, багульником, сфагнумом А4. Ряд заканчивается расположенной у подножия «сосной по болоту» V бонитета А5; в покрове здесь преобладают сфагнум, пушица, клюква и другие крайние влаголюбы. Береза встречается в качестве примеси к сосне всюду, кроме сухого бора.
Почвы в этом профиле сменяются последовательно от так называемых «боровых песков» в сухом и свежем бору через песчано-подзолистые почвы во влажном и глееподзолистые в сыром бору до торфяно-глеевых почв сфагнового болота. Слой грубого гумуса типично развит во влажном бору. В сыром бору он принимает торфянистый характер и увеличивается до 10—30 см, разрастаясь далее в мощный торфяной слой сфагнового болота. Все эти изменения сопровождаются приближением уровня грунтовых вод к поверхности почвы.
Особенности борового комплекса ярко отражены в сезонной динамике увлажнения отдельных его звеньев. Ранней весной почвы сухого и свежего бора достаточно увлажнены, но малые запасы почвенной влаги (низкая влагоемкость песка) быстро расходуются. Прирост древесины в этих борах, особенно в сухом, зависит от летних осадков. Влажный, сырой и заболоченный боры мало зависят от летних осадков и реже страдают от засух; более того, прирост насаждений влажного и сырою боров в средние по увлажнению годы обычно бывает выше, чем в годы с обильными осадками. Во второй половине вегетационного периода вследствие падения уровня грунтовых вод во влажных и сырых борах улучшается аэрация, а вместе с ней и другие почвенные условия, благодаря чему период сезонного роста по диаметру у них более растянутый.
Лишь в особо засушливые годы, сопровождаемые значительным снижением уровня грунтовых вод, прирост во влажных А3, сырых А4 и заболоченных А5 борах терпит больший ущерб, чем даже в свежих А2 и сухих А1 и А0 борах, так как эти последние лучше приспособлены к перенесению острой засухи. Кроме того, в каждом из типов боров, как это показано О. Г. Каппером (1938), в процессе естественного отбора возникают экотипы разной засухоустойчивости и гигрофильности. Так, в сухих борах преобладают сосны нестройного роста с сизой хвоей, ширококронные, с плакучими ветвями; в свежих и более влажных борах преобладают стройные узкокронные сосны с зеленой хвоей. Сосне по болоту также свойственны свои характерные формы.
Ход роста сосны в свежем и особенно в сухом бору связан с более ранней и резкой кульминацией прироста; во влажном и сыром борах кривые хода роста плавные. Более долговечны насаждения свежего бора (до 200 лет); недолговечны влажный и сырой бор, где деревья благодаря поверхностному характеру корневых систем заканчивают жизнь ветровалом в 100—120 лет.
С отмеченными биологическими особенностями насаждений связана различная эффективность рубок ухода. Более значительные степени изреживания эффективны в сухих и свежих борах; влажные и сырые боры после сильных изреживаний уменьшают прирост, так как при этом заболачивается почва. Смолопроизводительность сосны самая высокая в свежем и сухом бору; влажный и сырой боры менее эффективны для подсочки.
Наивысшие показатели механических свойств древесины сосны присущи свежему бору, в обе стороны от которого качество древесины ухудшается (А. Б. Жуков, 1931).
Естественное возобновление в борах также определяется ведущей ролью увлажнения. Наибольшее количество первичных вредителей-насекомых свойственно сухому бору; в сухих и свежих борах приобретает остроту вопрос борьбы с хрущами, подкорным клопом, побеговьюнами и другими насекомыми, находящими для себя оптимум в этих условиях. Во влажном и сыром бору иной состав вредной фауны и грибных паразитов, а в случае общих вредителей — иной ход заболеваний и дигрессии насаждений.
Боровой комплекс является результатом разных степеней увлажнения. Влажность играет ведущую роль в выработке экологических и лесохозяйственных особенностей разных звеньев борового комплекса. Однако при подробном его рассмотрении нередко делают вывод, что существенной причиной его разнообразия является не только меняющееся увлажнение, но и некоторые другие факторы и условия (например, рельеф, световой и тепловой режимы, почвенная аэрация и т. п.), которые играют при этом не меньшую роль, чем влага. Действительно, боровой комплекс является отражением рельефа, перераспределившего увлажнение, точнее — уровней грунтовых вод. Кроме того, на его отрезке, начиная с влажного бора и кончая «сосной по болоту», существенное влияние оказывает недостаток почвенной аэрации, отражением которого является падение бонитетов сосны от II до V. Несомненной является также существенная разница в световых условиях роста под пологом леса и тепловом режиме почвы. Наибольшее количество света и тепла получают кроны, стволы деревьев и напочвенный покров в слабо сомкнутых насаждениях сухого бора и «сосны по болоту».
Почвы сухого бора — самые бедные органическим веществом и наиболее сухие, поэтому они очень теплые; по мере нарастания влажности и накопления на поверхности органического вещества почвы становятся холоднее. Торфяно-глеевые почвы «сосны по болоту» — типичные холодные почвы.
Процессы накопления и разложения лесной подстилки в разных звеньях борового ряда различные. Имеется существенная разница в количестве и составе почвенных микробов, в количестве и составе надземной и особенно почвенной фауны, а также и в других элементах среды различных типов бора. Кроме того, сами по себе градации увлажнения испытывают неодинаковые количественные сдвиги под влиянием расходования влаги насаждениями, а это изменяет первичное (исходное) состояние увлажнения в рассматриваемом ряду.
Но было бы грубой ошибкой забыть о ведущей экологической роли увлажнения. Ближайшее рассмотрение показывает, что по отношению к влажности все прочие факторы, выступающие в боровом ряду, играют подчиненную роль. Приведем доказательства.
Выше было установлено, что влияние рельефа следует рассматривать как пространственное перераспределение экологических факторов, т. е. как влияние разных уровней грунтовых вод. Следовательно, рельеф как бы первый дает нам указание на то, что ведущими экологическими факторами, разнообразящими боровой профиль, являются разные степени увлажнения. В рассматриваемом случае влажность выступает в качестве ведущей причины по отношению к почвенной аэрации. Сначала на отрезке профиля от сухого бора к влажному благодаря приближению к ризосфере уровня грунтовых вод (в маловлагоемких песчаных почвах, являющихся весьма существенным источником влаги) производительность насаждений возрастает. Чем ближе находятся грунтовые воды к поверхности почвы, тем большее количество корней их достигает, тем полней обеспечено водоснабжение сосняков, особенно ценное в засуху. Но начиная с влажного бора, уровень грунтовых вод настолько высок, что в весенний период грунтовые воды, подымаясь по капиллярам, достигают поверхности почвы, и корни испытывают угнетение от недостатка воздуха.
Корни сосны могут развиваться лишь в верхней части зоны постоянного капиллярного увлажнения, лежащей выше зеркала грунтовых вод. Однако здесь они угнетены, часто отмирают и поэтому встречаются в небольшом количестве
В этом случае увлажнение и аэрация вступают в антогонизм и влага становится косвенным ограничителем производительности леса, сохраняя в то же время и свое прямое влияние на лес (водоснабжение корешков). Влияние нарастающего увлажнения становится подавляющим, и это вызывает резкие качественные изменения в лесорастительном эффекте. Качественные преобразования сопровождаются в дальнейшем иными количественными изменениями, если в отрезке профиля от сухого до влажного бора (А1—А2) шло увеличение производительности, то в отрезке влажного сырого бора и болота (А3—А5) производительность падает.
Некоторые лесоводы игнорируют специфическое влияние избыточного увлажнения, относя его к «косвенно действующим факторам» и противопоставляя ему «прямо действующий фактор аэрации». Между тем почвенная аэрация и анаэробиозис влияют на растения и прямо и косвенно. Недостаток воздуха в почве влияет на рост леса в значительной степени косвенным путем, сокращая ризосферу, «уплощая» корневые системы, заставляя их развиваться исключительно в поверхностных горизонтах почвы, т. е. там, где аэрация удовлетворительна и анаэробиозис устанавливается на относительно короткие периоды.
Следовательно, ближайшей причиной падения бонитета и других изменений в ряду А3—А5 выступает нарастающий почвенный анаэробиозис. Он влияет на растения как прямо, так и косвенно и целиком подчиняется фактору увлажнения.
Нетрудно показать, что различия в тепловом режиме почв, в световом и тепловом режиме атмосферы под пологом леса, в ходе накопления и разложения лесной подстилки, в микробиологической деятельности почв, да и самые типы почв борового ряда являются результатом различного увлажнения. Гораздо более сложные связи существуют, например, между составом надземной фауны и увлажнением, но и в этом случае увлажнение является решающим, ибо разный состав насаждений и их строение — есть посредствующие звенья, через которые влажность почвы (уровень грунтовых вод) определила в конечном счете также и состав фауны.
Таким образом, увлажнение является определяющим во всех различиях между отдельными звеньями (типами леса) рассмотренного нами борового ряда. Следовательно, его разнообразие обязано как в целом, так и в каждой из его сторон ведущей роли фактора влажности. Не может быть сомнений в том, что это экологический ряд последовательного гигрогенного замещения в его самом типичном выражении.
Экологический ряд (комплекс) боров разнороден по увлажнению, но однороден по почвенному плодородию, по количеству пищи растений, по химическим условиям плодородия.
Термин «химические условия плодородия» часто заменяют более кратким — «химическое плодородие». Г. Н. Высоцкий для обозначения того же понятия применял термин «трофность» (от греч. «трофос» — пища). Этот термин наиболее краток, применим как к почве, так и к растению. Трофность почвы — количество пищи растений в почве (химическое плодородие); трофность растения — его требовательность к почве, к количеству почвенной пищи, к химическому плодородию почвы.
Доказательством этой однородности является общность для всего борового ряда таких существенных показателей (индикаторов) бедных почв, как сосна и береза. Последняя, впрочем, отсутствует в сухом бору, но по другой причине — недостатку влаги. Существенными индикаторами предельно низкого плодородия во всем профиле являются низкие бонитеты, что становится вполне ясным, когда сравниваются боры с суборями эквивалентного увлажнения, Общим для всех звеньев борового ядра индикатором низкого почвенного плодородия является также отсутствие в нем растений средней или высокой требовательности к плодородию почвы.
Но если игнорировать лесорастительный эффект, пренебречь индикаторами и оценивать условия местообитания только по почвам, то изотрофность борового ряда не представится очевидной. Несмотря на однородность материнской почвенной породы (кварцевого песка), на пей образуются различные типы почв.
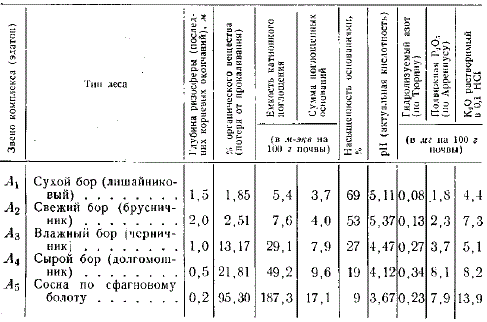
Данные анализа верхних почвенных горизонтов борового ряда
В таблице приведены данные анализа почв борового ряда в Межигорском лесничестве Первомайского лесхоза Киевской обл. Мы ограничились показателями верхнего горизонта (0—20 см), который в песчаных почвах сосредоточивает основной запас органических и подвижных минеральных веществ. Из таблицы видно, что более влажные и заболоченные почвы богаче органическим веществом и при большей емкости поглощения абсолютно богаче и основаниями. Вследствие замедленного разложения органического отпада актуальная кислотность и ненасыщенность почв основаниями от сухого бора к болоту увеличивается. В том же направлении увеличивается количество гумуса, поглощенных оснований, гидролизуемого азота, а также подвижных соединений фосфорной кислоты и калия. Однако из-за того, что ризосфера при этом значительно сокращается, плодородие почв всех звеньев ряда приводится к одинаковому уровню олиготрофных сосняков.
Во всех боровых почвах констатировано ничтожно малое количество бактерий, а в почвах сухого бора — также и грибов, представленных преимущественно плесенями. Их много только в лесной подстилке и в самых верхних горизонтах торфа более увлажненных звеньев ряда. Но и здесь, особенно в отрезке А3—А5, деятельность микрофлоры подавлена анаэробиозисом, кислой реакцией и малым количеством электролитов в почвенных растворах.
Иными словами, низкое плодородие борового ряда тесно связано с малым количеством и слабой активностью микрофлоры. После заселения боровых почв березой повышается микробиологическая активность почв, усиливается разложение органического вещества, вследствие чего в отдельных случаях боровые участки переходят в разряд суборевых.
На основе приведенных данных следует сделать вывод, что почва является сложным комплексом среды и наиболее точное и полное суждение о ее плодородии можно составить лишь в результате изучения ее лесорастительного эффекта. Рассмотрение лесорастительных свойств почвы в приведенном примере убеждает нас в отсутствии существенной разницы в почвенном плодородии (трофности) между отдельными звеньями борового комплекса. Следовательно, мы имеем право признать звенья борового ряда изотрофными, т. е. равноценными по плодородию, хотя и резко различными по увлажнению.