Рассмотрев факты, доказывающие эволюцию видов, набросав общую картину последней, Дарвин переходит к анализу ее причин. Этот анализ открывается замечательными мыслями, которые показывают глубину его.
Дарвин подчеркивает, что самый факт изменчивости и образования даже резко обозначившихся разновидностей, «мало поможет нам в понимании того, каким образом виды возникают в природе».
Следовательно, учение об изменчивости не вскрывает причин эволюции. Ключ к объяснению эволюции Дарвин видит в выяснении причин, по которым виды, эволюируя, в то же время приспособляются к новым условиям существования, и притом во всех жизненно-главных чертах морфо-физиологической организации. Эволюция видов, по Дарвину, совершается через эволюцию их приспособлений, т. е. носит приспособительный характер.
Следовательно, причинное объяснение эволюционного преобразования (исторической трансформации вида) может быть достигнуто через причинное объяснение истории его приспособляемости к определенным условиям существования.
Здесь происходит тот же процесс, что и при эволюции культурных форм. Последние эволюируют, приспособляясь к интересам человека. По аналогии с культурными формами можно принять, что приспособленность естественных видов — также результат отбора наиболее приспособленных, но производится он не человеком, а самими условиями среды.
Обратимся к некоторым примерам, которые покажут явление приспособленности организмов к условиям среды.
1. Приспособления или адаптации организмов к условиям жизни (относительная целесообразность) проявляются прежде всего в морфофизиологической организации живых форм. Ближайшим образом адаптация выражается в соответствии морфофизиологической организации любой живой формы условиям ее местообитания и образа жизни.
Чтобы показать этот замечательный факт, остановимся на организации дельфинов. Предварительно напомним, что млекопитающие — типичные наземные позвоночные. Дельфины, как и все киты, без сомнения происходят от сухопутных предков. Тем не менее, они полностью утратили связи с сушей и могут существовать только в воде. В связи с этим они достигли высокого совершенства приспособлений к водному образу жизни.
Тело дельфинов приобрело форму торпеды. Все выдающиеся части, мешающие скольжению в воде, либо полностью исчезли (наружное ухо), либо «убраны» в гладкий, обтекаемый корпус тела (млечные железы самок, наружные мужские половые органы). Эпителий кожи приобрел как бы полированную поверхность — свойство, увеличивающее легкость скольжения в воде. Образовался жировой спинной плавник. Передние конечности превращены в ласты, соответствующие грудным плавникам рыб. Задние конечности исчезли, уступив свою функцию жировому хвостовому плавнику. Эта замена конечностей хвостовым плавником усовершенствовала движение. Толкающий, движущий, локомоторный орган отнесен далеко назад, на «корму» торпеды. Это приспособление настолько же совершеннее гребущих лап, насколько винтовой пароход, с его кормовым винтом, совершеннее колесного. Работая хвостовым плавником, дельфин мчится по воде, как моторная лодка, по временам выбрасывая от сильного хода всю переднюю часть тела в надводное положение и развивая скорость до 50 км в час. Внутренняя его организация также сильно изменена. Под кожей развился мощный слой жира, уменьшающего удельный вес животного и играющего роль изолятора, препятствующего чрезмерной отдаче тепла через лишенную шерсти кожу. Утрачена не только ушная раковина, но и полностью зарос наружный слуховой канал. Замечательные преобразования претерпели ротовая полость и носоглотка. Зубы утратили обычную для млекопитающих форму. Нет диференцировки на резцы, клыки, премоляры и моляры. Все зубы приобрели приближенно одинаковую форму гарпунных зубов, приспособленных не для жевания, а для захватывания и удерживания рыб. Дельфин, не прожевывая, заглатывает свою добычу вместе с водой. При таких условиях любое наземное млекопитающее легко захлебнулось бы водой, попадающей в дыхательное горло. У дельфинов этого не бывает, так как гортань у них приподнята и прижата к внутренним носовым отверстиям (хоанам). Таким образом, дыхание осуществляется только через носовую полость (у дельфинов непарную), тогда как через ротовую полость дыхание не осуществимо. Это устройство гарантирует от попадания воды в гортань через рот. Слюнные железы исчезли, они не нужны, поскольку зубы потеряли жевательную функцию. В этой же связи язык утратил подвижность и стал похож на язык рыб. Носовые отверстия соединились в одно непарное, переместившееся далеко назад, на лоб, и прикрытое спереди щитком, препятствующим проникновению воды в носовую полость.

Приспособления к водному образу жизни у дельфинов. 1 — черноморский дельфин Phocaena relicla (Из Абеля): 2 — дельфин (Delphinus delphis), плывущий за парохода. На быстром ходу корпус выброшен из воды (По фото, из Illustration); 3 — медианный разрез через голову дельфина (Phocaena communis). Череп залит пунктиром. 1 — мозг, 2 — язык, 3 — гортань, 4 — пищевод (Из Гентшеля)
Процесс приспособления к водному образу жизни воздействовал также на особенности размножения дельфинов. Как правило, у млекопитающих при рождении детеныш появляется на свет головой вперед. Случаи, когда детеныш рождается в обратном положении, аномальны и затрудняют роды. По данным Слепцова (1940) у черноморского дельфина Delphinus delphis имеются обратные отношения. Дельфины рождаются хвостовым концом вперед. Это положение при рождении имеет адаптивное значение. Первым актом новорожденного млекопитающего является вздох. Вздох наполняет легкие воздухом и служит первой ступенью к новой, постэмбриональной жизни. Рождение дельфина головой вперед повело бы к тому, что голова в критический момент перехода от утробной к внеутробной жизни погрузилась бы в воду. Первый вздох — введение в постэмбриональное существование — был бы не возможен. Напротив, — рождение хвостом вперед оставляет голову некоторое время над водой. Этого момента достаточно, чтобы легкие наполнились воздухом. Уже через час после рождения новорожденный дельфин способен самостоятельно следовать за матерью. Отметим, наконец, что осмотическое давление крови у дельфинов близко к таковому у морских рыб (костистых).
Все эти преобразования не нарушили, однако, основных признаков, характерных для класса млекопитающих. Два затылочных мыщелка в черепе, пятипалый скелет грудного ласта, строение позвоночника и переднего пояса конечностей, наличие диафрагмы, левая дута аорты, постоянная температура тела, безъядерность эритроцитов, поразительно высокое развитие полушарий переднего мозга, разобщенность выводных путей пищеварительной и мочеполовой системы, вынашивание плода в матке, развитие типичных околоплодных оболочек, наличие усов (вибриссов) у эмбрионов, закладка на ранних стадиях развития двух пар конечностей, кормление молодых молоком, — все эти и многие другие признаки свидетельствуют о принадлежности дельфинов к плацентарным млекопитающим.
Таким образом, дельфины — настоящие млекопитающие, однако изменившиеся, в соответствии с водным образом жизни, т. е. приспособившиеся к нему. Приспособленность формы выступает здесь как объективное явление органической природы.
Подобные же явления наблюдаются и у растений, морфологические признаки которых претерпевают преобразования, зависящие от условий среды и достигающие порой поразительного совершенства.
Особенно интересные приспособления наблюдаются в строении так называемых энтомофильных, т. е. опыляемых насекомыми цветов.
Дарвин, специально изучавший процесс опыления у орхидных, описывает его в частности для одной британской орхидеи Orchis mascula.
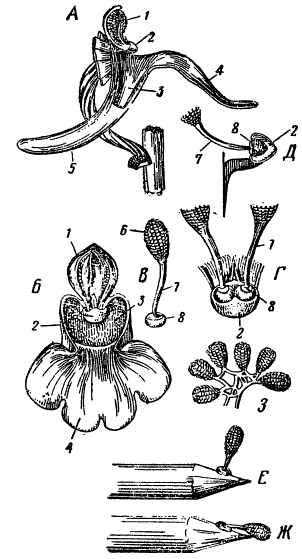
Орхидея Orchis mascula. 1 — двухгнездный пыльник, 2 — клювик. 3 — рыльце, 4 — губа. 5 — нектарник, 6 — пыльцевая масса, 7 — хвостик поллиния, 8 — липкие диски поллиния: А — сбоку, Б — спереди, В — один поллиний (видны пакетики пыльцы), Г — поллиний спереди. Д — разрез через клювик, Е — пыльцевая масса, только что прикрепившаяся, Ж — она же после окончания процесса опускания, 9 — пакетики пыльцы, связанные эластичными нитями (По Дарвину)
Цветок этой орхидеи снабжен длинной шпорцей, которая служит нектарником. При рассматривании цветка спереди обнаруживается, что над входом в нектарник находится рыльце, а под рыльцем небольшой клювик, служащий основанием для тычинок. Последние снабжены пыльниками в виде особых гнезд, наполненных пыльцой и называемых поллиниями. Поллинии возвышаются над рыльцем, а несущие их две тычинки опираются на свои широкие основания, имеющие форму дисков. Подошва последних клейкая и, в свою очередь, опирается на внутреннее углубление клювика.
Когда пчела садится на нижнюю губу цветка и пробирается отсюда в шпорцу (нектарник), она касается головой клювика. Внешняя стенка клювика немедленно разрывается, вследствие чего обнажаются клейкие основания дисков, на которых сидят обе тычинки с их поллиниями. Оба диска приклеиваются при этом к голове насекомого и, следовательно, обе тычинки с поллиниями уносятся им. Вскоре они, завядая, наклоняются вперед, и когда пчела проникает в следующий цветок, то поллинии оказываются в таком положении, что непосредственно соприкасаются с рыльцем и, следовательно, опыляют его.
Предположим теперь, что в цветке орхидеи имеются два рыльца, разделенных срединной перегородкой. При этом условии тычинки описанной организации оказались бы мало пригодными, так как была бы возможность, что поллинии, принесенные насекомым, прикоснутся не к рыльцам, а к перегородке между ними. Очевидно, при наличии двух рылец было бы полезно устройство, гарантирующее одновременное опыление обоих рыльцев. Эго имеется у орхидеи Orchis (anacantis) pyramidalis. Цветок этой орхидеи с двумя рыльцами, между которыми расположен клювик, опущенный в самый канал нектарника. Обе тычинки сидят не на двух дисках, как у О. mascula, а на одном общем диске, нижняя поверхность которого клейкая, как у О. mascula. Прикосновение бабочки (пытающейся ввести хоботок в нектарник) к клювику вызывает горизонтальный разрыв его передней стенки, вследствие чего обнажается клейкая поверхность диска. Последний имеет седловидную форму, причем хоботок бабочки, направляемый в нектарник, проходит как раз под седловидным диском, который и прилипает к хоботку. Мало того, под действием воздуха диск все более загибает свои края и плотно охватывает хоботок, почему возникает впечатление, будто бы диск пронзен хоботком. Когда диск охватывает таким образом хоботок, то обе тычинки, стоявшие первоначально на диске под небольшим углом друг к другу, взаимно все больше отклоняются, как два разделенных пальца. В таком положении они и уносятся бабочкой. Угол, образованный двумя тычинками, соответствует расстоянию между обоими рыльцами, и когда бабочка, несущая тычинки, проникает в новый цветок, обе поллинии прикасаются к двум им противостоящим рыльцам. Таким образом, достигается одновременное опыление обоих рылец. Дарвин с полным основанием рассматривает описанное устройство цветка, как одно из самых изящных приспособлений к опылению.
Действительно, описанные приспособления представляют удивительный механизм, все части которого тонко приспособлены к акту опыления при помощи насекомого.
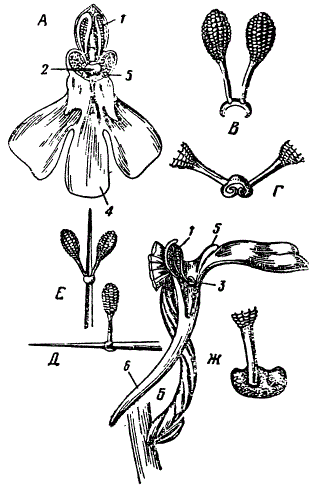
Орхидея Orchis pyramidalis. 1 — пыльник, 2 и 3— клювик, 4 — губа, 5 — направляющая пластинка на губе, 6 — нектарник, А — спереди, Б — сбоку, В — два поллиния, прикрепленные к седлообразному диску, Г — диск, не охвативший какого-либо предмета после сокращения, Д — поллинии, удаленные посредством введения иглы в нектарник; игла охвачена седельцем вследствие сокращения, Е — те же поллинии после второго движения и последующего опускания, Ж — диск сверху, насильственно сплющенный. Видно углубление, вызывающее второе движение поллиния (По Дарвину)
Ограничимся приведенными примерами, показывающими, что организация растений и животных многосторонне адаптирована к условиям среды. Эта адаптированность организации, по Дарвину, достигается естественным отбором.