Одна из характерных особенностей развития лесной растительности и почвообразования под пологом леса — ежегодное возвращение с опадающей листвой (хвоей), плодами, отмирающими ветвями и усыхающими в процессе самоизреживания отдельными деревьями части взятых из почвы элементов питания, при удержании на много лет другой части элементов в продолжающем развиваться древостое. Освобождаемые в результате минерализации отмерших частей лесной растительности элементы питания могут вновь потребляться развивающимся древостоем. В результате этого одно и то же количество элементов питания за время существования данного лесонасаждения многократно совершает круговое движение: почва — деревья — опад — лесная подстилка — почва. В данном случае в опад включаются и деревья, отмирающие в процессе самоизреживания древостоя. Этот процесс получил название малого биологического круговорота элементов.
В ходе малого биологического круговорота элементов следует различать три ветви:
- газообмен между листовой поверхностью растений и атмосферным воздухом;
- поглощение листовой поверхностью растений углерода из атмосферного воздуха, а корневыми системами растений из почвы азота, зольных элементов и воды, закрепление их в растительном организме, последующее поступление в почву после отмирания растений или их частей, разложение и освобождение заключенных в них элементов;
- прижизненное выделение корневыми системами некоторых элементов непосредственно в почву, поступление органического вещества и элементов питания за счет отмирания корней и корневых волосков.
Изучение первой ветви биологического круговорота составляет область физиологии растений. Основные работы в области лесного почвоведения были направлены на изучение второй и третьей ветвей биологического круговорота, а именно, обмена элементов между растительностью и почвой. Изучение третьей ветви — непосредственное взаимодействие и обмен элементов между корневыми системами и почвой, началось только в самые последние годы.
Практическая важность потребления различными древесными породами элементов питания из почвы и возвращения в почву взятых из нее элементов с опадом давно привлекали внимание исследователей к изучению отдельных частей биологического круговорота в лесу. Во второй половине XIX века ряд работ в этом направлении был опубликован в Германии и Франции. Эти работы Гейера и Фонхаузена, Вебера, Шрёдера, Раманна и Анри. Особенно большое значение имели исследования Эбермайера.
Исследования Эбермайера были проведены в лесах Баварии. Им было заложено 87 пробных площадей, на которых был организован учет массы лесной подстилки в продолжение 8 лет и определение годичного прироста в насаждениях бука, пихты, сосны, дуба, лиственницы и ели. Собранные образцы подстилки и древесины были подвергнуты химическому анализу в целях определения содержания азота и зольных элементов.
Проведенные работы выявили значительное накопление в лесной подстилке элементов питания. Из этого следовала ценность лесной подстилки как естественного удобрения, поддерживающего плодородие лесных почв. Поэтому сгребание лесной подстилки и последующий вывоз ее из леса обедняет лесную почву биологически важными элементами. Таким образом была доказана вредность практиковавшегося ранее удаления подстилки из леса.
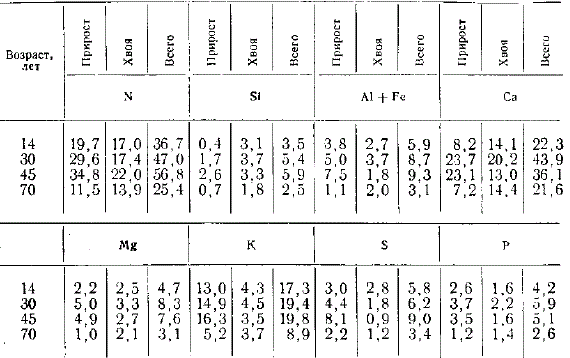
На основании полученных аналитических материалов Эбермайер попытался подойти к установлению количества элементов, извлекаемых лесной растительностью из почвы, которые возвращаются с опадом и удерживаются в составе древостоя, т. е. анализу биологического круговорота элементов питания в лесу. Для сосны и бука им были найдены следующие показатели.
Содержание элементов питания в лесной подстилке, кг/га в год
| Наименование показателей | Сосна | Бук | ||||||
| N | Ca | K | P | N | Ca | K | P | |
| Берется из почвы | 45 | 29 | 7 | 5 | 50 | 96 | 15 | 13 |
| Возвращается в почву | 35 | 19 | 5 | 4 | 40 | 82 | 10 | 3 |
| Удерживается в древостое | 10 | 10 | 1 | 1 | 10 | 14 | 5 | 10 |
Полученные материалы выявили существенные различия потребления и круговорота элементов питания в хвойных и широколиственных древостоях: большее потребление широколиственными породами (бук) по сравнению с хвойными (сосна); более интенсивный круговорот элементов и большее поступление в почву с опадом оснований, особенно кальция, в широколиственных лесах по сравнению с хвойными. Важна была и популяризация самой идеи круговорота элементов в лесу.
В настоящее время, когда прошло более 80 лет после опубликования Эбермайером его исследований, мы можем легко найти в них ряд методических несовершенств. Для своего времени и многих последующих лет исследования Эбермайера были крупным шагом в деле изучения взаимоотношений между лесом и почвой. Высоко оценивая вклад Эбермайера в учение о лесе, все же приходится отметить наиболее существенные методические недочеты в его исследованиях.
Эбермайер исследовал содержание элементов в листве (хвое) и древесине спелых и приспевающих древостоев. Вычисляя биологический круговорот элементов, он исходил из неверного предположения, что древостой во всех классах возраста берет из почвы и возвращает ей одинаковое количество азота и зольных элементов. В действительности же прирост органической массы с возрастом леса изменяется, а следовательно, меняется количество элементов питания, поглощаемых лесом из почвы. Кроме того, с возрастом леса изменяется количество деревьев, приходящихся на единицу площади, происходит самоизреживание древостоя, что неизбежно влияет на весь ход биологического круговорота. Эта сторона Эбермайером совсем не была учтена. Равным образом не был учтен и анализ корневых систем. Учет опада был проведен очень несовершенно, поэтому количественные характеристики слагаемых биологического круговорота оказались учтенными не полностью.
Исходя из более высокого процентного содержания азота и зольных элементов в листве (хвое) по сравнению с процентным содержанием в стволе и других частях дерева, Эбермайер сделал вывод, что основное количество поглощаемых из почвы элементов питания расходуется на создание листовой массы и в очень небольшой части — на годичный прирост древесины. При этом не было принято в расчет различие в массе ежегодно создаваемой листвы (хвои) и древесины. На основании сделанного Эбермайером вывода в дальнейшем усилия многих исследователей были в основном направлены на химический анализ листвы (хвои) без одновременного учета и исследования других частей дерева.
После опубликования работ Эбермайера несколько десятилетий не появлялось в печати равноценных работ по изучению биологического круговорота в лесу. Интерес исследователей, как уже было сказано, был направлен исключительно на химический анализ листвы, в меньшей мере — лесных подстилок. В результате был накоплен значительный аналитический материал, характеризующий содержание зольных элементов, а иногда и азота, в листве (хвое) различных древесных пород. Эти исследования, как правило, проводили без одновременного учета массы листвы (хвои). Подобные данные были у нас опубликованы Н. Н. Степановым. в зарубежной литературе Бродфут и Пире.
В несравненно меньшем масштабе велись исследования по учету опада, массы листвы, хвои, плодов, веток, ежегодно сбрасываемых древесным пологом на поверхность почвы. Поскольку учет массы опада не сопровождался его химическим анализом, то не могло быть получено данных о размере поступления в почву с опадом азота и зольных элементов.
Отдельные попытки дать химический анализ всех частей дерева принадлежат Коунслеру, Е. И. Парфеновой, Л. И. Троицкому, Н. Н. Дзенс-Литовской. Недостатком первых трех работ было изучение отдельных произвольно выбранных деревьев, без стремления подойти к освещению потребления и круговорота элементов.
Работа Н. Н. Дзенс-Литовской значительно ценнее, но она повторила ошибку Эбермайера, построив все свое исследование на анализе деревьев лишь одного возраста (около 40 лет).
В этих исследованиях содержание элементов зольного питания в стволе и ветвях делили на число лет дерева, произвольно принимая, что ежегодно дерево брало из почвы одинаковое количество элементов питания, независимо от возраста дерева.
Поэтому данные Н. Н. Дзенс-Литовской могут быть использованы лишь для сравнительной характеристики содержания зольных элементов в листве и древесине отдельных пород лесостепи. Крупным недостатком всех этих работ было отсутствие данных по определению азота.
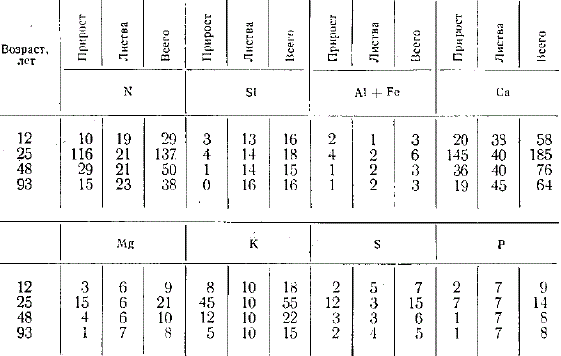
Новый этап в изучении биологического круговорота в лесу создали исследования кафедры почвоведения Московского университета (1944—1961 гг.). В основу этой новой серии исследований в отличие от предыдущих работ был положен динамический принцип, т. е. определение элементов, слагающих биологический круговорот по классам возраста древостоя.
Закладка пробных площадей и выбор модельных деревьев были произведены в соответствии с методикой, принятой при таксационных работах. На основании перечета и измерения диаметра и высоты деревьев выбирали характерные модели для данного древостоя. В сложных насаждениях брали модели сопутствующих пород, подроста и подлеска. Деревья валили, обрубали ветви, обрывали листву (хвою), раскапывали корневые системы. Путем взвешивания определяли отдельно массу листвы (хвои), мелких и крупных ветвей, ствола, крупных и мелких корней. В сумме получали вес всего дерева. Из всех частей дерева брали средние пробы для химического анализа. Для сбора опада были поставлены специальные ящики площадью 1 м2 каждый, по 10 ящиков на пробную площадь. Результаты определения веса отдельных частей дерева и данные химических анализов выражали на высушенное при 100° вещество. Модельные деревья в целях получения сравнимых данных брали в пределах одного срока, а именно: в августе, начале сентября (до листопада). Одновременно брали для анализа образцы травяно-мохового покрова и почвы.
Количество элементов питания, ежегодно вовлекаемых в биологический круговорот, определяли по сумме веществ, идущих ежегодно на создание массы листвы (хвои) и на годичный прирост ствола, ветвей, корней дерева. При этом принимали во внимание многодетность хвои и соответственно расчет производили по ежегодно создаваемой части общей массы хвои. Возвращение в почву элементов определяли по сумме их поступления с опадом и отмирающими в процессе самоизреживания деревьями. При этом расчет на отмирающие деревья вели по среднему модельному дереву подчиненного яруса, полагая, что в первую очередь отмирают деревья, отставшие в росте. Разность между этими двумя величинами характеризовала количество элементов, удерживаемых в развивающемся древостое.
Исследования позволили выявить следующие общие для всех изученных древесных пород закономерности. Процентное содержание азота и зольных элементов в различных частях дерева с возрастом изменяется очень мало. Колебания лежат в пределах точности исследования и не обнаруживают уловимых закономерностей. Исключения составляют деревца моложе 5 лет. Это открывает возможность, во-первых, пользоваться средними данными анализа деревьев разных классов возраста, а во-вторых, при дальнейших исследованиях ограничиться химическим анализом деревьев не всех классов возраста, что значительно облегчит и удешевит работу.
Далее были выявлены следующие общие различия в содержании азота и зольных элементов в различных частях дерева. Наиболее богаты всеми элементами листья (хвоя), затем (в нисходящем порядке) мелкие ветви, крупные ветви и ствол, древесина которого содержит наименьшее количество азота и зольных элементов. Мелкие корни по содержанию азота и зольных элементов близки к мелким ветвям, а крупные корни — к крупным ветвям.
Из всех элементов листья больше всего содержат азота, меньше кальция и еще меньше калия. Ветви, корни и ствол больше всего содержат кальция, затем идут азот и калий. В наибольших количествах во всех частях дерева содержатся кальций, азот, калий. Обращает внимание значительное содержание серы, близкое к содержанию фосфора. Алюминия содержится значительно больше, чем железа. Кремний в большинстве исследованных материалов присутствует в заметных количествах, значительно превышающих содержание алюминия. Железо, марганец и натрий были обнаружены в очень небольших количествах.
Наряду с отмеченными общими закономерностями в содержании азота и зольных элементов были обнаружены и существенные различия между исследованными древесными породами, обусловленные неодинаковой избирательной поглотительной способностью в отношении элементов питания.
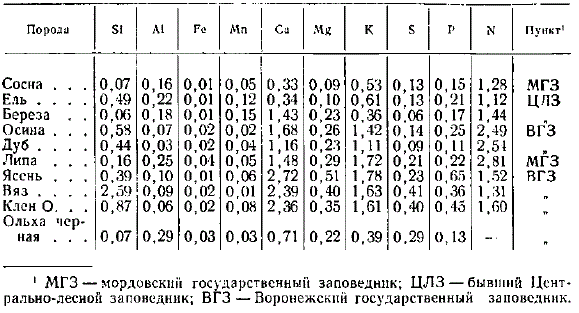
Содержание в среднем азота и зольных элементов к листьях (хвои) различных древесных пород (в % на высушенное при 100° вещество)
Из таблицы можно сделать следующие выводы о содержании азота и зольных элементов в листве (хвое). Наиболее богаты азотом листья липы (2,81%), далее в нисходящем порядке идут дуб и осина, значительно беднее азотом листья остальных пород, особенно хвоя сосны и ели (около 1%). Кальцием наиболее богаты листья ясеня (2,72%), близки к нему листья вяза и клена остролистного, далее в нисходящем порядке идут листья осины (1,68%), липы (1,48%), березы (1,43%) и дуба (1,16%). Меньше всего азота содержат хвойные породы (0,33%). Из других особенностей следует отметить высокое содержание кремния в листьях вяза (2,59%), значительно меньше в листьях клена остролистного (0,87%), осины (0,58%), в хвое ели (0,49%), листьях дуба (0,44%). Остальные породы еще беднее им. Наименьшее содержание кремния найдено в хвое сосны (0,07%) и листьях березы (0,06%). Фосфором наиболее богаты листья ясеня (0,65%) и клена остролистного (0,45%). Серы больше всего содержится в листьях вяза (0,41%), клена (0,40%). Обращает внимание низкое содержание в листьях дуба фосфора (0,11%) и серы (0,09%). Наибольшее количество алюминия содержится в листьях липы (0,25%).
Дуб, ясень, вяз, клен остролистный были взяты с одной пробной площади, следовательно, росли в одинаковых условиях. В том же Воронежском заповеднике и на близких почвах была взята осина. Таким образом, отмеченные различия в содержании азота и зольных элементов обусловлены биологическими особенностями сравниваемых древесных пород. Равным образом ель и береза были взяты в близких почвенных условиях в Центральном лесном заповеднике.
Древесина ствола по сравнению с другими частями деревьев наиболее бедна азотом и зольными элементами. Поэтому различия между древесными породами в содержании названных элементов становятся менее заметными. Все же древесина вяза и клена отчетливо выделяется более высоким, а хвойных и березы — более низким содержанием кальция.
Различия в содержании указанных элементов в листве древесных пород не полностью повторяются при сопоставлении с их содержанием в древесине ствола. Так, например, листья березы содержат больше кальция, чем листья дуба, а ствол березы отличается его небольшим содержанием, приближаясь в этом отношении к хвойным. Следовательно, береза, давая относительно много кальция с опадающей листвой, сравнительно мало его расходует на годичный прирост древесины.
Ветви мелкие и крупные по содержанию азота и зольных элементов занимают соответственно промежуточное положение между листвой (хвоей) и древесиной ствола.
В данных анализа корней обращает внимание низкое содержание кремния в корнях ели. Корни липы, осины, дуба выделяются более высоким содержанием кальция, в меньшей степени калия. Особое внимание привлекают корни черной ольхи. В них по содержанию элементов первое место занимает азот, второе — кальций (на некоторых почвах — алюминий), третье — калий. Накопленные аналитические данные могут служить материалом для суждения о том, в какой мере содержание азота и зольных элементов в различных частях дерева представляет характерное свойство отдельных древесных пород и в какой мере на него влияют почвенные условия. Из материалов других исследователей оказалось возможным использовать данные В. Н. Мины по дубнякам Теллерманова леса и Н. Н. Дзенс-Литовской по Савальскому лесничеству Воронежской области. Как и в других случаях, влияние почвенных условий наиболее рельефно отразилось на содержании азота и зольных элементов в листве (хвое). В хвойных лесах имеет значение также степень сомкнутости крон.
Различная сомкнутость крон в редком и густом 35-летнем сосновом насаждении (Эйтннген, «Лесоводство»)
Сравнение содержания названных элементов в хвое сосны из сосняков брусничного, черничного и липнякового, несмотря на различия в почвенных условиях (слабодерновая среднеподзолистая песчаная почва, мощный песчаный подзол слабо оглеенный, бурая лесная супесчаная почва), показывает близкое их содержание. Все же можно отметить меньшее содержание кальция и магния в хвое сосны, выросшей на подзоле в сосняке-черничнике (0,15% и 0,06%) по сравнению с двумя другими типами сосняков. (0,33 и 0,09%), а также меньшее содержание алюминия (0,12%) на бурой лесной почве по сравнению с более кислыми почвами подзолистого типа (0,16%). В общем можно сделать вывод, что в данном случае различие в почвенных условиях мало отразилось на содержании элементов питания в хвое сосны. Более резко отличались анализы хвои сосны из Урды. Следует отметить в хвое этой сосны более низкое содержание азота, фосфора и особенно калия и серы при соответственно повышенном содержании кремния, алюминия, кальция и магния.
Более существенные различия обнаружены в составе хвои ели. Особенно выделяется ельник ручейниковый, где принос с верховодкой элементов зольного питания обусловливает более обильное обеспечение кальцием и калием. Было найдено следующее содержание кальция в хвое ели: в ельнике-зеленомошнике на суглинистом подзоле — 0,48; в ельнике сложном на дерново-подзолистой суглинистой почве — 0,75; в ельнике ручейниковом — 1,35%. Сходная зависимость содержания в хвое кальция от почвенных условий наблюдалась и при исследовании ельников Архангельской обл. Хвоя ельника ручейникового богаче также калием и кремнием.
Листья березы, выросшей на сравнительно богатых перегноем, но более легких по механическому составу бурых лесных супесчаных почвах МГЗ богаче азотом, но беднее кальцием, чем листья березы ЦЛЗ, выросшей на дерново-подзолистых суглинистых почвах.
Листья осины из осинника осоково-снытевого на серо-бурых лесных супесчаных почвах ВГЗ богаче азотом, калием, фосфором, чем листья из осинника широкотравного на дерново-подзолистых суглинистых почвах ЦЛЗ. Листья из осинника лещиново-осокового на оподзоленном супесчаном черноземе Савалы отличаются от осинника на серо-бурых лесных супесчаных почвах ВГЗ меньшим содержанием кальция и алюминия, при большем содержании калия и близком других элементов. Меньшая величина содержания алюминия в анализах Н. Н. Дзенс-Литовской по Савальскому лесничеству, по-видимому, обусловлена несовершенством метода анализа.
В данном случае не принималось во внимание, что при высоком содержании фосфорной кислоты она с полуторными окислами осаждается не полностью. При вычитании из суммы полуторных окислов содержания фосфорной кислоты, определенной отдельно, получаются заниженные показания для алюминия. Сказанное относится и к другим определениям содержания алюминия в работе Н. Н. Дзенс-Литовской.
Листья липы из двух разных мест произрастания не обнаружили существенных различий, хотя в одном случае были взяты листья из липняка снытево-осокового МГЗ, а в другом из ельника сложного ЦЛЗ, отличающихся по почвенным условиям.
Листья дуба из Воронежского заповедника и Савальского лесхоза, развитые на близких почвах супесчаного механического состава, мало различаются по содержанию зольных элементов. Можно отметить лишь более низкое содержание фосфора и серы при более высоком содержании кремния в листве дуба Воронежского заповедника. Листья дуба из Теллермана, выросшие на более богатых известью почвах, отличаются несколько более высоким содержанием кальция и магния.
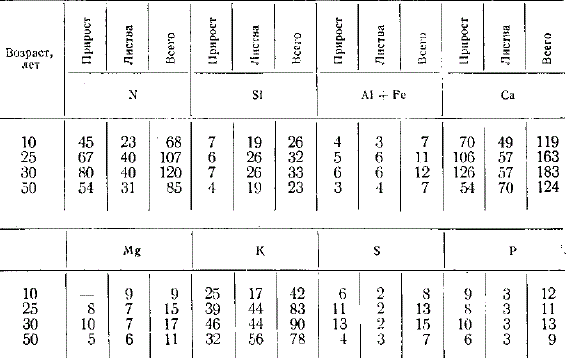
В разные стадии развития древесные породы потребляют неодинаковое количество азота и зольных элементов. В качестве общей закономерности следует отметить возрастание потребления азота и зольных элементов с возрастом дерева, но у разных пород это идет в неодинаковой степени.
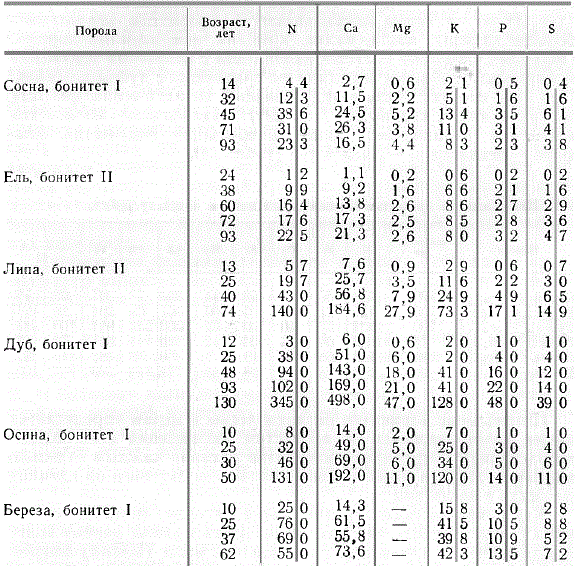
Потребление азота и главнейших зольных элементов одним деревом в зависимости от возраста, г/год
Можно отметить следующие особенности потребления главнейших элементов питания отдельными деревьями, образующими в совокупности лес, Хвойные деревья берут из почвы значительно меньше элементов питания, чем лиственные. Это особенно четко выступает, если сравнить потребление в одном возрасте. Например, в 90—100 лет одно дерево дуба берет ежегодно кальция 169 г, а дерево сосны в том же возрасте 17 г, ели 21 г; азота соответственно берут одно дерево дуба 102 г, сосны 23 г, ели 23 г. Из исследованных хвойных пород сосна потребляет больше элементов питания, чем ель. Поскольку сосна была первого бонитета, а ель второго, то указанное различие в известной мере может быть объяснено неодинаковым классом бонитета. Хвойные породы больше всего берут из почвы азот, второе место по потреблению занимает кальций, третье калий, далее идут потребляемые в близких количествах магний, сера, фосфор. Для сосны следует отметить значительно большее потребление азота по сравнению с кальцием, ель обнаруживает меньшее различие в потреблении обоих названных элементов.
Широколиственные породы и осина берут из почвы больше кальция, чем азота. Третье место по потреблению занимает калий, далее идут остальные элементы. Обращает внимание неодинаковая скорость потребления элементов питания с возрастом. У дуба возрастание потребления с возрастом идет постепенно. Резкое увеличение потребления элементов питания дубом в 130 лет обусловлено сильным осветлением древостоя, вызванным выборочными рубками, что привело к усиленному разрастанию оставшихся дубов. Этот крутой подъем потребления нельзя рассматривать как закономерный природный процесс.
Следует отметить резкое увеличение потребления элементов питания липой после 40 лет и осиной после 30 лет. Дуб выделяется среди остальных пород относительно большим потреблением серы и фосфора. В возрасте около 50 лет одно дерево дуба берет кальция 143 г, а осины 192; азота соответственно 94 и 131 г; калия 41 и 120 г и т. д.
По абсолютным размерам накопления азота и зольных элементов в одном дереве с возрастом сравниваемые породы существенно отличаются друг от друга. Широколиственные породы (дуб, липа) и осина накапливают с возрастом значительно больше кальция, азота, калия, кремния, чем хвойные породы. Обращает внимание большее накопление с возрастом алюминия и марганца хвойными породами по сравнению с дубом и осиной. При этом следует иметь в виду меньший возраст осины (50 лет) по сравнению с хвойными (100 лет). Наибольшие количества алюминия накапливает липа, наибольшие количества фосфора — осина, серы — липа и дуб.
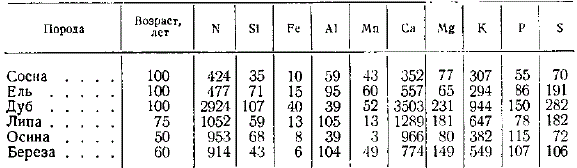
Накопление в древостое элементов питания к возрасту рубки, кг/га
Потребление элементов питания лесом в целом определяется двумя изменяющимися с возрастом насаждения величинами: возрастанием потребления элементов питания каждым отдельно взятым деревом и уменьшением количества деревьев на единицу площади с возрастом.
Как показали исследования, масса ежегодно создаваемой листвы (хвои) существенно возрастает лишь в самые первые годы, а в дальнейшем изменяется сравнительно мало. Поэтому колебания в потреблении элементов литания лесом в целом зависят в большей степени от величины годичного прироста всей массы органического вещества деревьев и кустарников, растущих на данной площади. В данном случае имеет значение именно общая масса создаваемого органического вещества: ствола, ветвей, корней, т. е. физиологический годичный прирост, а не объемный прирост, учитываемый таксаторами.
Если сопоставить величины физиологического годичного прироста, т, е. всей массы органического вещества для исследованных типов насаждений, то выявятся следующие закономерности. Для всех исследованных насаждений имеет место постепенное повышение годичного прироста с возрастом до времени достижения наивысшей точки, характерной для данного типа леса. После этого начинается сначала крутое, а затем более постепенное снижение прироста.
Период наибольшего прироста органического вещества в разных типах леса наступает не одновременно; в сосняке-брусничнике в возрасте 30—50 лет; ельнике зеленомошно-кисличном около 40 лет; дубняке осоково-снытевом и липняке снытево-осоковом около 25 лет; в осиннике около 30 лет. Сопоставление с таблицами хода роста, показывающими объемный прирост, позволяет установить совпадение наибольшего годичного прироста для сосняков, дубняков и осинников. У липняков и ельников было обнаружено наступление наибольшего физиологического прироста, учитываемого по всей массе синтезируемого органического вещества, на 15—20 лет раньше наибольшей величины прироста, показанного в нормальных таблицах хода роста, которые учитывают объемный прирост. Годичный прирост по всей массе синтезируемого органического вещества, конечно, всегда больше товарного прироста, показанного в нормальных таблицах хода роста.
Наибольшая величина годичного прироста всей массы органического вещества, согласно исследованиям кафедры почвоведения Московского университета, составляет в т/га для липняка 11, сосняка 15, ельника 20, осинника 24, дубняка 36. Из сказанного видно, что в возрасте 25—30 лет в дубняках и осинниках синтезируется особенно большая масса органического вещества, что, в свою очередь, должно сопровождаться потреблением большого количества элементов питания. В зависимости от местных условий и истории развития насаждений от приведенных величин неизбежны отклонения.
Далее встает вопрос о соотношении расхода элементов питания на ежегодное создание листовой (хвоевой) массы и на годичный прирост органического вещества. Эбермайер, Раманн и другие исследователи полагали, что несравненно большая часть элементов питания, берущихся древесными породами из почвы, идет на создание листвы (хвои) и сравнительно небольшая — на годичный прирост древесины. На этом основании было принято судить об относительном потреблении зольных элементов различными древесными породами по данным химического анализа листвы (хвои), а именно, по процентному содержанию в них золы.
В настоящее время вопрос о соотношении расхода элементов питания на ежегодное создание ассимиляционного аппарата и прирост древесины (ствол, ветви, корпи) может быть рассмотрен на основе экспериментальных материалов кафедры почвоведения Московского университета.
Потребление азота и зольных элементов на создание хвоевой массы и годичного прироста в сосняке, кг/га в год
Исследования в сосняке брусничном показали, что примерно до 50-летнего возраста сосновый древостой на физиологический годичный прирост расходует больше азота, кальция, калия, магния, серы и фосфора, чем на ежегодное создание хвои. Исключение составляет кремний, который в большем количестве поступает в хвою. Для сосняка 70 лет было найдено заметное сокращение расхода элементов на годичный прирост; на первое место для всех исследованных элементов вышел расход на создание хвоевой массы.
Несколько иначе складывается рассматриваемое соотношение в дубняках. Здесь очень рельефно выступает больший расход азота. Расход всех элементов питания на создание листового аппарата довольно постоянен, он очень мало колеблется с возрастом дубового леса. Решающее влияние имеет размер годичного прироста. Соотношение складывается таким образом, что в возрасте жердняка расход азота, кальция, магния и серы на годичный прирост намного превышает их потребление на построение листовой массы. В остальные классы возраста расход элементов питания на годичный прирост заметно меньше, чем на создание листвы. Особое место и здесь занимает кремний. Он во все возрасты почти полностью расходуется на листовую массу. Его потребление листвой во все возрасты значительно выше расхода на прирост.
Потребление азота и зольных элементов на создание листовой массы и годичного прироста дубняком, кг/га в год
В осиннике в возрасте до 70 лет включительно потребление азота и фосфора на прирост превышает его расход па создание листовой массы. Кальций и сера в большем количестве расходуются на прирост до 30 лет. Калий идет в примерно одинаковых количествах на прирост и листву до 30 лет, а затем его расход на прирост существенно сокращается. Для кремния сохраняется та же закономерность, что была отмечена для других древесных пород. Кремний поступает главным образом в листву и лишь в очень небольшом количестве удерживается в составе прироста.
Потребление азота и зольных элементов на создание листовой массы и годичного прироста в осиннике, кг/га в год
Приведенные материалы показывают ошибочность представления о том, что большая часть элементов, извлекаемых древесной растительностью из почвы, расходуется па создание листовой (хвоевой) массы, а не на годичный прирост (в физиологическом понимании). У насаждений разного породного состава, в разном возрасте и для различных элементов это соотношение изменяется в значительных пределах: в одних случаях больше расход на листву, в других — на прирост. Из этого следует, что по содержанию рассматриваемых элементов в листве еще нельзя судить об их потреблении на прирост, а следовательно, и о потреблении древостоем в целом.
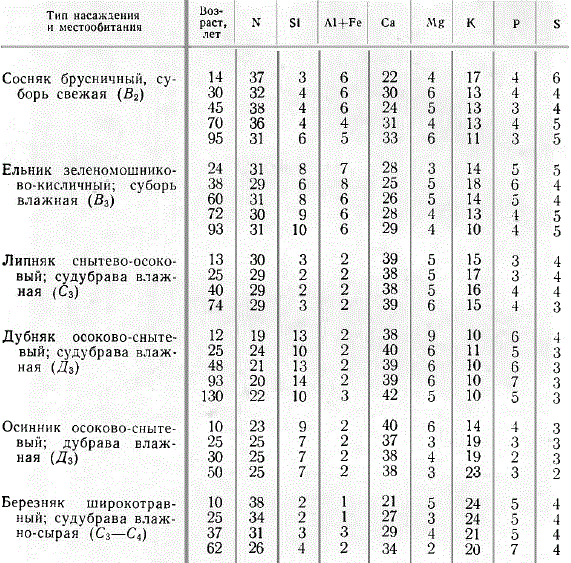
Наименьшее количество элементов питания (из числа исследованных) берет из почвы сосновый лес; далее в порядке возрастания потребления идут ельник, дубняк, липняк, осинник, березняк. Иллюстрацией могут служить величины наибольшего потребления перечисленными древостоями элементов питания (в сумме) в кг/га: сосняк 62, ельник 73, дубняк 133, липняк 162, осинник 258, березняк 517. Смешанные хвойно-широколиственные леса берут из почвы больше элементов питания, чем чистые хвойные. С повышением продуктивности древостоя возрастает потребление элементов питания. Приведенные выше величины суммарного потребления получены для древостоев I и отчасти II классов бонитета.
Размер потребления элементов питания изменяется с возрастом древостоя. В этом отношении можно наметить три группы типов леса. Первую группу составляют сосняки, ельники и дубняки, для которых найдено возрастание потребления к возрасту жердняка, т. е. к максимуму потребления всех элементов в 20—40 лет, после чего начинается снижение. Ко второй группе принадлежат липняки и осинники, у которых потребление элементов питания с возрастом изменяется сравнительно мало, период наибольшего потребления — около 30 лет, но выражен нерезко. Отдельно стоят березняки, образующие третью группу, для которой характерно очень высокое потребление всех элементов в возрасте около 10 лет, а затем прогрессирующее снижение потребления. Из общей схемы выпадет дубняк (130 лет), в котором природный процесс был нарушен около 20 лет тому назад выборочными рубками.
Хвойные (сосна, ель) и береза берут из почвы больше всего азота, на втором месте стоит кальций, на третьем калий, далее идут остальные элементы, потребляемые в значительно меньшем количестве, Широколиственные леса (дубняки, липняки) и осинники, в противоположность хвойным и березе, берут больше всего кальция, а потребление азота стоит на втором месте, третье место сохраняется за калием, остальные элементы потребляются в меньших количествах. Серу и фосфор большинство древостоев берет приблизительно в равных количествах, лишь дубняк и березняк выделяются несколько большим потреблением фосфора по сравнению с серой.
Существенно выделяются высоким потреблением кремния дубняки и осинники. Для березняков характерно высокое вовлечение в биологический круговорот калия, значительно большее, чем у остальных древесных пород.
Если вычислить потребление элементов питания в относительных процентах, т. е, приняв их сумму за 100, то выявляется сравнительно малая изменяемость с возрастом соотношения между элементами питания, вовлекаемыми данным типом древостоя в биологический круговорот.
Относительное потребление азота и зольных элементов лесом (в % от общего потребления)
Результаты этих вычислений показывают, что в зависимости от класса возраста изменяется количество элементов, которые лес берет из почвы и вовлекает в биологический круговорот, но соотношение между элементами остается довольно постоянным. Лишь в отдельных случаях можно заметить более значительные отклонения, что может быть связано с различиями в соотношении между приростами отдельных частей лесообразующих пород.
Постоянство соотношения между вовлекаемыми в биологический круговорот элементами питания по классам возраста позволяет выяснить средние относительные проценты потребления элементов питания.
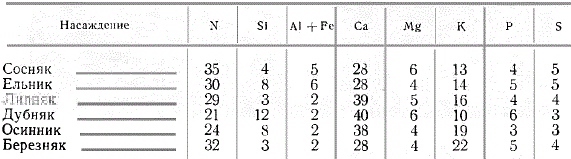
Среднее относительное потребление азота и зольных элементов различными древостоями (в % от общего потребления)
Относительное потребление кальция широколиственными породами (40—39%) и осинником (38%) более высокое, чем хвойными и березой (28%). Дубняк выделяется более низким относительным потреблением азота (21%), близок к нему осинник (24%). Наиболее высоким относительным потреблением азота выделяется сосняк (35%), далее в порядке убывания — березняк (32%), ельник (30%), липняк (29%). Следует отметить более высокое относительное потребление калия березняком (22%), осинником (19%) и наиболее низкое дубняком (10%). По высокому относительному потреблению кремния выделяются дубняк (12%), осинник и ельник (по 8%) по сравнению с остальными породами (4—3%). Хвойные выделяются большим потреблением алюминия и железа (в сумме).
Из сказанного следует общий вывод об устойчивости избирательной поглотительной способности как характерного биологического признака лесообразующих древесных пород. В процессе биологического круговорота лесная растительность берет из почвы и возвращает в нее элементы питания в том соотношении, которое наиболее благоприятно для развития лесной растительности. В результате смены поколений леса происходит обогащение верхних горизонтов почвы элементами питания в соотношении, необходимом для лесной растительности. Этим повышаются лесорастительные свойства почвы и подготавливаются условия для смены древостоями, более требовательными к почвенным условиям.
Ежегодно вместе с опадающей листвой (хвоей), ветками, плодами почва под лесом получает значительное количество органического вещества и заключенных в нем элементов питания. Проведенные исследования показали, что количество опадающей листвы (хвои), ветвей, плодов и т. д., в зависимости от возраста и полноты насаждений, колеблется в следующих пределах в т/га на высушенное при 100° вещество:
| Сосняки чистые | 1,3—2,5 |
| Сосняки липняковые | 2,8—3,4 |
| Ельники чистые | 1,9—2,0 |
| Ельники сложные | 2,2—3,2 |
| Березняки | 2,7—3,2 |
| Осинники | 2,8—4,9 |
| Липняки | 2,0—3,5 |
| Дубняки | 3,1—4,5 |
Из приведенных данных следует, что чистые хвойные насаждения дают наименьшее количество опада (1,3—2,5 т/га), несколько больше дают хвойно-широколиственные, березняки и липняки (2,2—3,5 т/га), наибольшее — дубняки и осинники (2,8—5). Следует отмстить значительное колебание в поступлении опада по годам, что связано с влиянием метеорологических условий на жизнь леса. Многолетние наблюдения за опадом в дубравах Воронежского заповедника дали колебания для дубняка в возрасте около 100 лет от 2,8 до 5,5 г/га; дубняка около 50 лет от 3,3 до 4,9 r/га; дубняка около 50 лет от 3,3 до 4,9 т/га и т. д.
В зависимости от массы и качественного состава опада почва получает неодинаковое количество элементов питания.
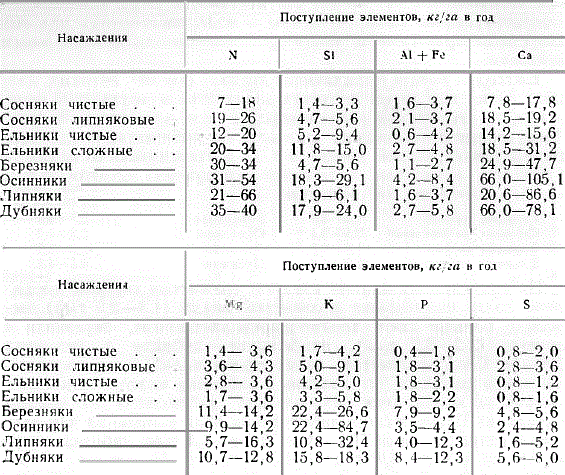
Поступление азота и зольных элементов с опадом в лесах разного породного состава
Резко выделяются чистые хвойные насаждения, опад которых приносит наименьшее количество почти всех элементов. В хвойно-широколиственных лесах опад заметно богаче азотом и кальцием, что связано с участием широколиственных пород. Еще богаче элементами питания опад широколиственных лесов и осинников. Липняки, дубняки и осинники дают с опадом большее количество азота, кальция, калия, фосфора, серы. Следует отметить значительное содержание в опаде березняков азота, фосфора и серы. Кальция больше всего поступает с опадом в осинниках, затем в порядке убывания идут липняки, дубняки и березняки. Наблюдается высокое поступление с опадом в дубняках и осинниках кремния, больше чем в лесах другого породного состава.
В ельнике, липняке и осиннике поступление элементов питания с опадом по классам возраста довольно равномерное в течение исследованного периода. Исключением служат молодняки, когда опада поступает значительно меньше. Для некоторых элементов наблюдается небольшое увеличение поступления с опадом при переходе к старшим классам возраста. В сосняках и дубняках возврат азота и зольных элементов с опадом постепенно увеличивается и достигает наибольшей величины во II—III классах возраста, а затем снижается. Для дубняка отмечено повторное увеличение поступления этих элементов с опадом при переходе от V к VII классу возраста. Как уже было сказано, это обусловлено выборочными рубками, которые привели к разрастанию крон первого яруса, развитию подроста и подлеска.
В насаждениях, где не проводят регулярно рубок ухода, значительное количество органического вещества, азота и зольных элементов поступает в почву с сухостоем, образующимся в процессе самоизреживания древостоя. В большинстве случаев поступление в почву азота и зольных элементов с сухостоем меньше поступления с опадом. Лишь в период формирования древостоя с отмирающими в процессе самоизреживания деревьями поступает больше мертвого органического вещества и заключенных в нем элементов питания.
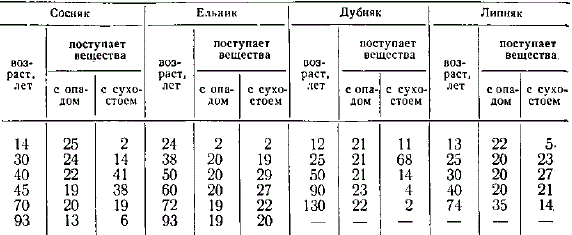
Поступление в почву мертвого органического вещества с опадом и сухостоем, ц/га
Мертвые органические вещества с сухостоем в насаждениях различного породного состава поступают не одновременно, соответственно изменяется и время наибольшего поступления в почву с сухостоем азота и зольных элементов. Например, в сосняке период наибольшего поступления сухостоя приходится на 40—45 лет, в ельнике 50—60, дубняке около 25 лет, липняке около 30 лет.
Несколько иначе происходит самоизреживание в осиннике. Здесь количество сухостоя с возрастом увеличивается, параллельно идет и увеличение массы опада.
| Возраст древостоя, лет | 10 | 25 | 50 |
| Сухостой, ц/га | 3 | 25 | 78 |
| Опад, ц/га | 21 | 30 | 33 |
При накоплении данных по большему количеству пробных площадей эти сроки могут быть изменены в зависимости от истории развития каждого насаждения.
Химический анализ стволов растущих и усохших деревьев не обнаружил существенной разницы. Поэтому расчет элементов, поступающих с сухостоем, не требует дополнительного химического анализа сухостоя.
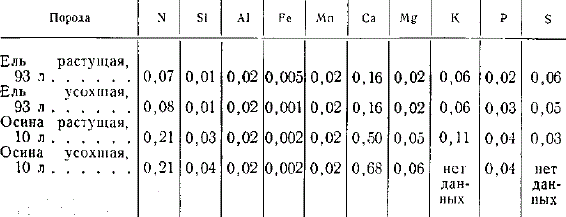
Содержание азота и зольных элементов в стволе растущих деревьев и сухостое (в % на высушенное при 100° вещество)
Если выразить количество элементов, возвращаемых почве с опадом и сухостоем, в процентах от потребления этих же элементов в том же возрасте, то выявится следующая закономерность. В начальный период развития большинства исследованных типов леса, т. е. первые 30—40 лет, возврат составляет около 40—60% от потребления соответствующих элементов. Остальная часть удерживается в составе древостоя. Лишь в единичных случаях отмечены отклонения за указанные пределы.
Поступление в почву азота и зольных элементов с опалом и сухостоем (в % от потребления)
Во второй период в приспевающих и спелых насаждениях в составе древостоя удерживается ежегодно значительно меньшая часть взятых из почвы элементов и возврат составляет 80—90% от потребления, лишь в единичных случаях спускаясь до 60%. Исключением служит дубовый лес, который уже в первом классе возраста возвращает почве около 70—80% от потребления.
В поведении отдельных элементов привлекает внимание высокий возврат кремния во всех классах возраста в дубняках (80—98%) и осинниках (71—92%) при соответственно низком проценте его удержания в составе текущего прироста.
В сложных многоярусных насаждениях, подобных дубравным, в биологическом круговороте, кроме главной породы, принимают участие сопутствующие породы, подрост и подлесок. В дубравах отмечено участие (кроме дуба) ясеня, вяза, ильма, клена остролистного, липы, лещины, клена тюлевого и татарского, груши, яблони и т. д. По данным проведенного исследования, наибольшее влияние сопутствующие породы оказывают в молодых насаждениях до приспевающих включительно. Особенно велико влияние сопутствующих пород в дубняке 50 лет, когда их участие по массе достигает 15%, к 100 годам оно падает до 1%.
Влияние спутников дуба отражается на расходе азота и зольных элементов, потребляемых для создания листовой массы и годичного прироста, а также па поступлении в почву названных элементов с опадом и сухостоем. В приведенных показателях встретились трудности с учетом массы отмирающих деревьев сопутствующих пород, подроста и кустарников подлеска. В дальнейшем повторный перечет на постоянных пробных площадях поможет восполнить этот пробел. В приведенных ранее данных биологического круговорота в дубняке внесены необходимые поправки на участие в круговороте других деревесных пород, кроме дуба.
С необходимостью учета влияния прочих древесных и кустарниковых пород пришлось столкнуться и при изучении биологического круговорота в сосняках и ельниках сложных. Участие широколиственных пород (дуба, липы, лещины) повышало интенсивность биологического круговорота, особенно — кальция, азота и калия. Заметное влияние этих пород начинало сказываться с третьего класса возраста, когда вследствие осветления древостоя под пологом хвойного леса получают возможность развиваться широколиственные породы.
Далее следует остановиться на влиянии травяно-мохового покрова на биологический круговорот в лесу. Полученные аналитические материалы показывают существенные различия в содержании азота и зольных элементов между зелеными мхами (Pleurozium Schreberi, Hylocomium proliferum, Dicranum undulatum), лесными кустарничками (брусникой, черникой) и дубравным широкотравьем.

Содержание азота и зольных элементов в травяномоховом покрове лесов
Наиболее существенные различия были обнаружены в содержании кальция и калия. В зеленых мхах, кислице, дубравном широкотравье калий количественно преобладает над кальцием, что характерно для травянистой растительности. В чернике и бруснике, наоборот, калия содержится значительно меньше, чем кальция. В этом отношении брусника и черника приближаются к древесным породам. Наиболее высоким содержанием калия и кальция выделяется характерное растение дубравного широкотравья — сныть, оно богаче других и азотом. Наименьшее количество калия содержат брусника, черника и зеленые мхи. Осока и папоротник выделяются высоким содержанием кремния. Дубравное широкотравье богаче других растений алюминием. Корневые системы дубравного широкотравья по сравнению с надземной частью заметно беднее азотом, кальцием, калием, кремнием, магнием.
В покрове из зеленых мхов, как показывают данные пересчета на 1 га, содержится сравнительно небольшое количество всех элементов. Если учесть, что мхи, нарастая с одного конца, отмирают с другого, то следует вывод об их сравнительно небольшом непосредственном участии в биологическом круговороте. Еще меньшее количество всех элементов содержится в покрове из кислицы, создающей очень небольшую массу органического вещества. Поскольку покров из кислицы ежегодно полностью отмирает, то это определяет большее его влияние на биологический круговорот в лесу по сравнению с зеленомоховым. Брусника и черника, как многолетние растения, оказывают очень небольшое влияние на биологический круговорот.
Наиболее сильное влияние на биологический круговорот в лесу оказывает покров из дубравного широкотравья, который вовлекает в биологический круговорот значительное количество азота (до 26 кг/га), кальция (16,6 кг/га), калия (36 кг/га), кремния (7,1 кг/га) и т. д. Поэтому при исчислении биологического круговорота в дубняках, липняках, осинниках, а также хвойно-широколиственных лесах с покровом из дубравного широкотравья надлежит вводить соответствующую поправку на потребление элементов питания травянистой растительностью и возврат по мере отмирания травяного покрова. В наибольшей степени участие травяного покрова отразится на увеличении биологического круговорота калия. Это обусловлено тем, что травянистая растительность вовлекает в биологический круговорот больше всего калия, в то время как по количественному потреблению древесной растительностью калий занимает третье место (после кальция и азота). На почвах, недостаточно обеспеченных физиологически доступными соединениями калия, развитие травяного покрова может существенно влиять на питание древесных пород калием.
Определенное влияние на биологический круговорот в описываемых лесах оказывают лесные эфемероиды. Подснежник, гусиный лук, ветреница лютиковая, хохлатки, медуница начинают свое развитие еще под снегом. Уже в январе под покровом снега можно обнаружить зеленые ростки подснежников. После схода снега довольно быстро начинается массовое цветение эфемероидов. В это время деревья и кустарники, еще не одетые листвой, в большом количестве пропускают солнечные лучи, способствуя развитию покрова из эфемероидов.
Произведенные подсчеты показывают, что эфемероиды дубового леса только в надземной части содержат в кг/га: азота 4—9; калия 8—18; кальция 2—4 и остальные элементы в меньших количествах, Эфемероиды развиваются ранней весной, когда почву промывают талые снеговые воды и весенние дожди. Вовлекая в биологический круговорот калий, который особенно легко вымывается из мертвых растительных остатков, эфемероиды препятствуют его вымыванию. Если учесть, что в дубовом лесу с опадающей листвой ежегодно поступает в почву 16—18 кг/га калия, то значение его поглощения эфемероидами (8—18 кг/га) проявляется еще яснее.
А. Я. Орлов предлагает различать в корневой системе древесных пород три части, резко отличающиеся по своему участию в биологическом круговороте элементов:
- основные крупные скелетные корни, существующие в течение всего периода жизни дерева; накопленное в них вещество поступает в почву лишь после отмирания деревьев;
- мелкие проводящие корни, отмирающие и заменяющиеся новыми в процессе формирования и жизни корневой системы;
- сосущие корни — постоянно обновляющийся аппарат, служащий для поглощения питательных веществ и воды. Продолжительность жизни сосущих окончаний, вероятно, не превышает нескольких лет.
По участию в круговороте веществ первая часть корней аналогична стволу и крупным ветвям, вторая — мелким ветвям, опадающим в течение всей жизни дерева в связи с формированием кроны и очищением ствола от ветвей, и третья — листьям и хвое, которые периодически обновляются.
Наибольшее значение в обогащении почвы органическим веществом принадлежит отмирающим сосущим корням, которые сравнительно быстро поддаются минерализации, приводящей к освобождению заключенных в них элементов.
А. Я. Орлов согласен с И. В. Тюриным, что учет прижизненного отмирания корневых волосков и покровных тканей вряд ли осуществим. Кроме того, для большинства древесных пород СССР, кроме хвойных, этот источник поступления органического вещества имеет небольшое значение, так как корневые волоски у этих пород образуются лишь на ростовых корнях и общая их масса очень невелика.
По вопросу о продолжительности жизни сосущих корней в литературе нет единого мнения. Одни авторы полагают, что сосущие корни древесных пород существуют более одного вегетационного периода, другие, что они отмирают в тот же вегетационный период, в который образуются.
Наиболее подробные исследования были проведены В. А. Колесниковым по изучению корневых систем яблони. Он пришел к выводу, что образование новых корней и отмирание старых идет в течение всего вегетационного периода. По его наблюдениям отмирают не только корневые окончания, но, как он пишет, целые «корневые системки», понимая под этим боковые корни, имеющие несколько порядков корней. В. А. Колесников считает, что весной и летом преобладает нарастание корневых систем, а начиная с сентября и до весны следующего года происходит отмирание не только кончиков корней, но и целых «корневых системой» или, по его терминологии, «самоизреживание корневых систем». По наблюдению ряда авторов, после засушливого лета, при наступлении осенних дождей, имеет место второй период нарастания корней.
Интересная попытка количественного учета содержания живых и отмерших сосущих корней и других органических остатков в почве была предпринята А. Я. Орловым. В этих целях им была разработана специальная методика. Эта трудоемкая работа была проведена в культурах ели 25 и 50 лет на выщелоченном суглинистом черноземе (Моховской лесхоз Орловской обл.), что нельзя считать характерным местом произрастания ели.
А. Я. Орлов применил следующую методику. При помощи металлического каркаса сечением 100 см2 и высотой 10 см брали из почвенного профиля послойные образцы. Взятый объем почвы промывали на ситах диаметром отверстий 0,5 и 0,25 мм. Корневые окончания ели имеют диаметр 0,2—0,3 (0,4) мм. На ситах оставались «все живые и отмершие, не полностью разложившиеся корневые окончания, а также более толстые проводящие корни и другие органические остатки (тяжи мицелия, кусочки коры и т. д.). Сильно разложившиеся, легко распадающиеся на крупинки отмершие окончания корней проходили через отверстия сит».
Отделенные корни А. Я. Орлов исследовал под бинокулярной лупой (увеличение в 10 и 40 раз) и разделял визуально на фракции.

Количество живых и отмерших корней, гумуса и других органических веществ в толще почвы 0—200 см под культурами ели (по А. Я. Орлову), г/га сухого веса дерева
Исследование показало, что еловые культуры 25 лет имели в три раза более мощную систему мелких (диаметром > 0,5 мм) корней, чем культуры ели 50 лет. А. Я. Орлов объясняет это возрастными особенностями. Он полагает, что культуры ели 25 лет, находящиеся в возрасте наиболее интенсивного роста и при сильной загущенности, обусловили непропорционально мощное развитие сосущего аппарата. С этим объяснением можно было бы согласиться при наличии данных изучения корневых систем елей того же возраста, растущих в иных условиях.
Далее А. Я. Орлов берет сумму отмерших крупных и мелких корней, других отмерших органических остатков, тяжей мицелия, а также живых корневых окончаний и сопоставляет с массой подстилки. В еловом древостое 25 лет эта сумма составляет 23,3 г/га при массе подстилки 20 г/га; в древостое 50 лет соответственно 10,5 и 14,4 т/га, что свидетельствует о близости массы подстилки и корней. Однако из этого нельзя делать вывод об одинаковых темпах их накопления. Разложение мертвого органического вещества на поверхности почвы протекает значительно быстрее, чем в ее толще. Об этом свидетельствует нахождение в толще почвы ельника корней березы, которая росла здесь около 30 лет тому назад. Отмершие крупные корни и другие органические остатки (кусочки коры корней) представляют довольно устойчивые включения, медленно поддающиеся разложению. Заключенные в них элементы очень постепенно освобождаются и вступают в новый биологический круговорот.
Для определения количества ежегодно поступающей массы отмерших сосущих корней А. Я. Орлов приводит следующий расчет. В ельнике 50 лет корни диаметром 0,5 мм составляют 1,8 г/га. Из этой величины следует вычесть вес корней диаметром 0,3—0,5 мм, которые представляют многолетние образования. Следовательно, масса корневых окончаний и наиболее тонких проводящих корней (их всего 5—8%) составляет примерно 1,2 г/га. Если принять среднюю продолжительность жизни корневых окончаний в 2 года, то ежегодный отпад сосущих корней составит около 0,6 т/га. По исследованиям А. Я. Орлова, для разложения корневых окончаний до стадии крошения требуется около 3—5 лет. Если средняя продолжительность разложения 4 года, то ежегодная величина отпада сосущих корней около 0,5 т/га, т. е. величина, близкая к ежегодному отмиранию сосущих корней, что следует учитывать при изучении биологического круговорота. К сожалению, А. Я. Орловым не был произведен химический анализ выделенных из почвы корней. Поэтому осталось неизвестным участие содержащихся в отмирающих корнях зольных элементов в биологическом круговороте.
Более детальные работы по изучению участия деятельных корней в биологическом круговороте были проведены кафедрой почвоведения Московского университета. Исследования были проведены в дубравах Воронежского государственного заповедника. Бонитет дуба III, почва серо-бурая лесная песчаная. Легкий механический состав почвы облегчил препаровальную работу.
В описываемых исследованиях Е. М. Борискиной была применена методика А. Я. Орлова с небольшими изменениями.
На основании предыдущих исследований Л. Г. Богатовой было установлено, что основная масса деятельных корней дуба на этих почвах сосредоточена в верхнем слое. Образцы для исследования брали непосредственно под лесной подстилкой металлическим каркасом сечением 100 см2 и высотой 10 см, что соответствовало объему 1000 мл3. Взятый монолит делили на два слоя высотой по 5 см. Из каждой половины раздельно выбирали и учитывали массу сосущих корней. Подобные монолиты брали вокруг модельного дерева дуба на расстоянии 70 см от ствола (по наблюдениям А. Я. Орлова, количество сосущих корней не зависит от расстояния от ствола дерева). Монолиты были взяты три раза за сезон: в июне, июле и сентябре 1956 г., с 14 по 21 число каждого месяца. Повторность 5—6-кратная.
Е. М. Борискина предупреждает, что термин «сосущие корни» не вполне точен. Учитывались не только сосущие окончания, но и оси диаметром меньше 0,3 мм, на которых они располагаются. Большая часть осей и некоторые корневые окончания утратили функции всасывания.

Количество живых корней дна метро и менее 0,3 мм в слое почвы 0—10 см под дубом III бонитета в г на 1 л почвы (из работы Е. М. Борискиной)
Как видно из табл., среднее количество корней в монолите сечением 100 см2 и высотой 10 см в июне и июле составляло по 0,8 г, а в августе только 0,5 г, т. е. в июне и июле если и было отмирание сосущих корней, то оно компенсировалось нарастанием новых. В августе отмирание превысило нарастание, что привело к снижению общего содержания сосущих корней. Большая часть сосущих корней, около 90%, сосредоточена в слое 0—5 см, в следующем слое 5—10 см имеется очень небольшое их содержание. В горизонтах наибольшего сосредоточения средних и крупных корней дуба на глубине 20—40 см сосущих корней совсем не было обнаружено. Отходящие от крупных корней проводящие корни поднимаются вверх и дают сильное ветвление в гумусовом горизонте.
Подобное же распределение наблюдал В. Н. Мина в Теллермане на серых лесных суглинистых почвах, а также С. А. Самцевич и Т. Д. Катеринин на южных черноземах Украины.

Данные химического анализа сосущих корней дуба в сопоставлении с данными анализа других частей корневой системы (в % на высушенное при 100° вещество)
Данные анализа показали значительно более высокое содержание в сосущих корнях кремния, алюминия, железа и калия по сравнению с более крупными корнями.
Важно отметить, что отмирающие сосущие корни будут обогащать подвижными соединениями перечисленных элементов верхние горизонты почвы. Не исключена возможность переноса железа и алюминия из более глубоких горизонтов. К сожалению, за недостатком материала не удалось определить содержание азота.
При сопоставлении содержания зольных элементов в сосущих корнях и опаде отчетливо выявляется большое содержание в сосущих корнях железа и алюминия. Если принять продолжительность жизни сосущих корней два года, как допускает А. Я. Орлов, то и в этом случае им принадлежит более крупная роль в круговороте железа и алюминия. Кроме того, корни существенно влияют па круговорот кремния и калия.

Сопоставление содержания элементов в сосущих корнях и в опаде дубового леса, кг/га
Следующий вопрос, который был поставлен в план изучения роли корней древесных пород, было исследование прижизненной десорбции корнями некоторых элементов. Эти исследования в 1956 г. были проведены Е. М. Борискиной в дубняках и сосняках Воронежского заповедника и Н. А. Титовой в Беловежской Пуще (Белоруссия).
Для травянистых растений многими исследованиями установлено, что наряду с поглощением из почвы элементов питания имеет место десорбция или экзосмос корнями некоторых элементов обратно в почвы. Этот процесс имеет место преимущественно в конце вегетации. А. И. Ахромейко на основании исследования сеянцев ряда древесных пород пришел к выводу, что корневые выделения обнаруживают суточную периодичность. В начале вегетации в суточном ходе преобладает поглощение, а в конце — выделение некоторых элементов. Это совпадает с положением высказанным Н. А. Максимовым, что выделение некоторых элементов корневыми системами связано со старением растений. В это время из наземных частей некоторое количество элементов поступает в корни, а последними выделяется в почву. Со взрослыми деревьями подобных исследований не было произведено и не было разработано соответствующей методики.
В этой связи представляют интерес исследования Е. М. Борискиной при участии Л. Г. Богатовой, проведенные с 50—60-летними деревьями дуба и сосны. Методика была применена следующая.
Осторожно откапывали и освобождали от почвы боковой корень длиной 30—40 см с большим количеством сосущих корней и тщательно отмывали его от приставших частиц почвы. После этого корень погружали в склянку емкостью 1 л, наполненную колодезной водой, закрывали картонной крышкой с отверстиями для корня и стеклянной трубочки для аэрации. Сверху крышки помещали слой плотной бумаги и клеенки для более полной изоляции. Через 7—10 дней колодезную воду сменяли на дистиллированную. В некоторых опытах вместо дистиллированной воды применяли разбавленные растворы KNO3 и Ca(NO3)2. Имелись опасения, что при дистиллированной воде десорбция может быть обусловлена разностью концентраций. Через сосуды каждые 5 дней продували воздух для обеспечения достаточной аэрации. По прошествии месяца содержимое сосудов извлекалось для анализа, а сосуды наполнялись новыми порциями дистиллированной воды или указанными ранее разбавленными солевыми растворами. Следует отметить хорошее состояние корней во время опыта, которые даже давали прирост. Результаты получены следующие.
Корни древесных пород не только поглощают из почвы элементы питания, но и выделяют их в окружающую среду. Преобладание выделения над поглощением калия, кальция, магния и алюминия наблюдалось в конце августа — начале сентября. В остальные сроки имело место преобладание поглощения. Установлено подкисляющее влияние корневых выделений на почву, обусловленное углекислотой. Явления подкисления почвы корешками были констатированы и ранее в работах П. С. Погребняка. Е. М. Борискиной установлено, что в результате деятельности корней происходит повышение активной, обменной и гидролитической кислотности почвы и содержания в ней подвижного алюминия. Наряду с этим происходит повышение емкости поглощения катионов, которое обусловлено гуматной частью. Возрастание емкости поглощения происходит частью в результате повышения содержания обменных оснований. Это можно рассматривать как улучшение лесорастительных свойств почвы.

Данные химического анализа почвы прикорневой и внекорневой зоны (из работ Е. М. Борискиной)
Анализ почвы в прикорневой зоне и вне ее показывает, что в прикорневой зоне концентрация водородных ионов значительно выше, чем вне корневой зоны. Разница pH суспензий составляет значительную величину, Величина обменной и гидролитической кислотности в почве прикорневой зоны во всех случаях значительно превышает величину обменной и гидролитической кислотности внекорневой зоны. В некоторых случаях обменная кислотность почвы в прикорневой зоне в 5 раз и более превышает обменную кислотность почвы внекорневой зоны. Особенно велико это превышение в начале вегетации растений, когда жизнедеятельность корней проходит более интенсивно. В этот период обычно наблюдается и вспышка жизнедеятельности почвенных микроорганизмов. Обменная кислотность почвы почти полностью обусловлена подвижным алюминием, Содержание его в прикорневой зоне вдвое больше, чем во внекорневой.
Можно предполагать (Т. И. Евдокимова, Е. М. Борискина), что основная масса подвижного алюминия накапливается в прикорневой зоне в результате подкисления почвы корнями и растворения соединений алюминия, входящих в состав почвенных минералов. Определенную роль в накоплении в почве алюминия играют также и корневые выделения, в составе которых он обнаружен, и освобождение алюминия при разложении богатых этим элементом корневых систем растений.
В прикорневой зоне, несмотря на подкисление, не только не идет вытеснение катионов кальция и магния водородом, но происходит даже абсолютное увеличение их содержания в составе поглощающего комплекса. Источником обогащения почвы кальцием и магнием служат растительные остатки, а также корневые выделения.
В прикорневой зоне всегда происходит обогащение почвы органическим веществом в результате разложения отмерших тканей и сосущих корней.
Величина емкости поглощения показывает рост поглощающего комплекса в прикорневой зоне. Этот рост, очевидно, обусловлен в основном накоплением органического вещества, так как степень насыщенности основаниями вблизи корней ниже, чем во внекорневой зоне. Воздействие корневых систем на почву в значительной степени изменяет ее свойства.