Определение понятия «вид» представляется очень трудным делом.
Мы будем рассматривать вид как исторически сложившуюся совокупность организмов, занимающих определенный ареал обитания, и характеризующихся общностью происхождения, сходной системой приспособлений к уровням среды и воспроизведением в поколениях основных адаптивных черт и признаков.
Организмы одного вида обладают характерными для данного вида фенотипом и генотипом, отличными от таковых у организмов другого вида.
Вид занимает определенный ареал. Ареал распространения одних видов оказывается широким, и такие виды являются обычно политипическими — включают несколько географических рас, или подвидов. Другие виды имеют значительно более ограниченный ареал, они, как правило, не образуют географических рас и являются монотипическими. Особи, составляющие вид, не образуют собой неизменную однородную массу. Каждый организм вида, обладая общими и характерными чертами, имеет и свои индивидуальные генотипические особенности, представляющие в совокупности наследственную изменчивость вида, или, как иногда называют, «генофонд» вида.
Вид, представляя единую совокупность организмов, расчленяется на отдельные популяции. Популяцией называют совокупность свободно скрещивающихся особей одного вида, характеризующихся общностью местообитания и приспособления к данным условиям существования. Популяция складывается под влиянием условии существования на основе взаимодействия факторов наследственности, изменчивости, и отбора. Образование популяций является своеобразным способом «пригонки» вида к конкретным условиям его существования. Породы животных и сорта растений, создаваемые искусственным отбором, также представлены отдельными популяциями.
Процессы формирования популяций и их динамики составляют микроэволюцию. Возникновение новых видов начинается с дивергенции — расчленения вида на отдельные, нескрещивающиеся или изолированные группы организмов. Популяция является своего рода «кузницей», в которой естественный отбор творит новые формы.
В природе популяции каждого вида характеризуются генетическим разнообразием. Но мы часто этого не замечаем. Особи популяции и вида внешне кажутся нам относительно однообразными. Это относительное однообразие животных и растений, позволяющее систематикам относить животных и растения к определенным видам, подвидам, расам, создается естественным отбором. Отбор обеспечивает не только разнообразие, но и однообразие внутри вида.
Однако указанное однообразие относится лишь к основным типичным чертам, признакам и свойствам организмов данной популяции. Как только мы начнем детально генетически анализировать состав популяции, разлагая ее на отдельные линии, так сейчас же обнаружим огромную генотипическую изменчивость. При этом оказывается, что каждой популяции, в пределах которой особи длительное время скрещивались друг с другом, на ограниченной территории при данных климатических условиях присущ свой характер изменчивости.
Источниками наследственной изменчивости в популяции являются мутационная и комбинативная изменчивости. Действие генетических законов в популяции представляет предмет исследования генетики популяций.
В природе не существует двух организмов, абсолютно сходных между собой по генотипу. Зоологи или ботаники, изучающие биологические процессы у какой-либо группы особей, всегда имеют дело с наследственно разнородной группой организмов. Но поскольку ими изучается фенотип организмов, они вправе пренебрегать генетическим разнообразием своего материала.
Изучение популяции можно вести чисто описательным методом. В этом случае определяют фенотипическую характеристику форм популяции, ее биологические особенности, выясняют условия существования и взаимоотношение организмов, цепи питания, конкуренцию, динамику численности по годам и зависимость ее от разных факторов. Популяции обособляются и складываются в результате действия многих факторов: способа размножения, характера изменчивости, изменения численности особей, темпа и направления отбора, климатической, географической и физиологической изоляции. Основным из них является отбор признаков, обеспечивающих процесс воспроизведения поколений, т. е. размножение. Очевидно, что при разных способах размножения возникновение и поддержание популяций осуществляется разными путями, в чем можно убедиться, сравнивая популяции перекрестно оплодотворяющихся (аллогамных) и самооплодотворяющихся (автогамных) организмов.
Для существования популяции имеют первостепенное значение различные типы наследственной изменчивости: генные мутации, хромосомные перестройки и полиплоидия. Ненаследственные изменения могут играть лишь ограниченную роль. Организмы, генотипически различающиеся, например, по одному гену, морфологически могут не отличаться друг от друга, но обладать различными физиологическими особенностями (жизнеспособностью, длительностью развития, плодовитостью). Генетические методы дают возможность составить более полное представление о наследственных потенциях популяции, ее приспособительных особенностях и направлении эволюции.
Зачинателями изучения генетической структуры популяций надо признать селекционеров, ибо для того, чтобы выявить разнообразие генотипов в популяциях, необходимо было изолировать, подобрать для скрещивания отдельные пары родителей и затем исследовать их потомство в ряду поколений. Именно так поступали селекционеры, создававшие различные сорта и породы. Однако научные основы генетического изучения популяции могли быть заложены только после открытия Г. Менделя, установившего количественные закономерности наследования.
Первое научное изучение популяции, совместившее в себе генетические и статистические методы, было предпринято датским физиологом растений и генетиком В. Иоганнсеном. Его классическая работа «О наследовании в популяциях и чистых линиях», опубликованная в 1903 г., положила начало генетическому изучению популяций. Как часто бывает в науке, классическое открытие делается на элементарном, на первый взгляд, явлении и при использовании простой методики. Так и в данном случае В. Иоганнсен избрал объектом исследования популяции не перекрестноопыляющиеся, а самоопыляющиеся растения — ячмень, фасоль и горох. Методически это упрощало работу, поскольку каждую такую популяцию можно было легко разложить на группы потомков отдельных особей, т. е. произвести выделение отдельных, «чистых», линий. «Чистой линией, — писал он, — я называю индивиды, которые происходят от одного самоопыляющегося индивида. Отсюда ясно, что популяция абсолютных самоопылителей состоит только из чистых линий, индивиды которых в природе могут быть смешаны, но не могут быть затронуты скрещиванием».
В качестве признаков были взяты вес и размеры семян. Эти количественные признаки определяются действием многих генов, т. е. определяются полигенно, и в сильной степени подвержены изменчивости под влиянием факторов внешней среды — состава почвы, климата, способа посадки растений и т. п. Поэтому для установления характера их наследования необходимо использование математических методов анализа изменчивости.
По этим признакам имеется ярко выраженная модификационная, или паратипическая, изменчивость. О значении этой изменчивости для эволюции в биологии имелись различные точки зрения. Сторонники теории наследования приобретенных свойств считали, что изменения, вызванные воздействием факторов внешней среды, наследуются, передаются потомству. Противники этой теории отрицали наследование модификационных изменений. Решение этого спора в пользу последних имело принципиальное значение, так как отбор организмов по фенотипу без выявления наследственных потенций имел до этого широкое распространение в селекции и тормозил выведение пород животных и сортов растений.
Иоганнсен провел взвешивание семян одно сорта фасоли и построил вариационный ряд по этому показателю. Вес семян оказался изменчивым в пределах от 150 до 750 мг. В дальнейшем семена весом от 250 до 350 мг были высеяны отдельно от семян весом 550— 650 мг. С каждого выросшего растения вновь были взвешены семена. Так как фасоль является самоопыляющимся растением, то генотип семян с одного растения должен быть одинаковым, а у семян от разных растений могут быть генотипические различия. Поэтому тяжелые семена (550—650 мг) и легкие (250—350 мг), выбранные из сорта, представляющего популяцию, дали растения, семена которых значительно различались по весу. Средний вес семян на растениях, выросших из тяжелых семян, составил 518,7 мг, а на растениях, выросших из легких семян, — 443,4 мг. Этим было показано, что сорт-популяция фасоли состоит из генетически различных растений, каждое из которых может стать родоначальником чистой линии.
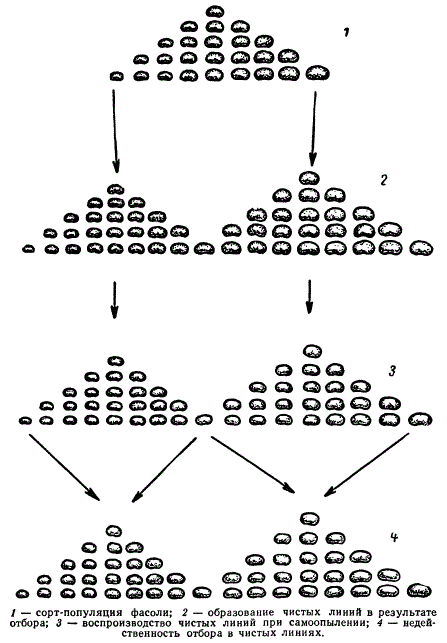
Схема, иллюстрирующая разложение популяции на чистые линии и недейственность отбора в них
На протяжении 6—7 поколений Иоганнсен отбирал также тяжелые и легкие семена с каждого растения в отдельности, т. е. производил отбор в пределах чистых линий. При таком отборе ни в одной линии не произошло сдвига в ряду поколений в сторону тяжелых или легких семян. Следовательно, изменчивость по весу семян внутри чистой линии была ненаследственной, модификационной.
В результате своих исследований Иоганнсен пришел к следующим выводам: 1) «отбор в популяции вызывает… больший или меньший сдвиг — в направлении отбора — того среднего признака, вокруг которого, флуктуируя, варьируют соответствующие индивиды» и 2) «внутри чистых линий регрессия (степень сходства признака потомства с материнским) была… полной; отбор внутри чистых линий не вызывал никакого смещения типа».
Как мы видим, популяция автогамных растений состоит из генотипически разнородных линий. Растения такой популяции не скрещиваются между собой и не обмениваются наследственной информацией. В этом случае существование популяции основывается на строгом естественном отборе линий определенного генотипа, на общности приспособительных механизмов к однотипным условиям внешней среды. Иначе говоря, изменение популяции автогамных растений и животных осуществляется отбором определенных наследственно различающихся линий и клонов, имеющих адаптивные преимущества.
При самооплодотворении отдельный организм может быть зачинателем новой расы, подвида и вида, а также сорта или породы. Например, новый сорт пшеницы может быть выведен от одного зерна, отобранного из популяции.
Однако, говоря о высокой гомозиготности в чистых линиях, следует иметь в виду, что даже чистые линии не могут быть абсолютно гомозиготными по следующим обстоятельствам. Во-первых, облигатных (абсолютных) самоопылителей-растений не существует. В популяциях самоопылителей, например пшениц, томатов и др., всегда с той или иной частотой встречаются растения с открытым цветением и перекрестным опылением. В силу этого между чистыми линиями в популяциях хотя и редко, но совершаются процессы скрещивания и соответственно обмена наследственной информацией. Во-вторых, у растений-самоопылителей встречаются мутации, препятствующие самоопылению (несовместимость). В-третьих, в чистых линиях самоопылителей даже за одно поколение возникает весьма ощутимое количество разнообразных мутаций, которые нарушают гомогенность чистой линии.
Вследствие указанных причин сорта растений-самоопылителей при воспроизведении в производстве могут утрачивать часть своих сортовых качеств и требуют постоянного контроля, на чем и основана необходимость сортообновления.
При вегетативном размножении агамных организмов, не имеющих полового процесса или вторично его утративших (некоторые простейшие, грибы, водоросли и др.), объектом отбора в популяции являются отдельные клоны. Генетическая целостность (интеграция) таких клонов в популяции является очень низкой в силу невозможности осуществления скрещиваний между особями разных клонов, но такие популяции, по-видимому, все же существуют в природе и поддерживаются отбором на основе симбиотических отношений разных генотипов.
У перекрестно оплодотворяющихся организмов в природе популяция формируется на основе свободного скрещивания разнополых особей с разными генотипами, т. е. на основе панмиксии. При этом наследственная структура следующего поколения воспроизводится на основе разнообразных сочетаний различных гамет при оплодотворении. Отсюда вытекает, что численность особей того или иного генотипа в каждом поколении будет определяться частотой встречаемости разных гамет, произведенных генотипически различными родительскими организмами. Это значит, что признаки и свойства сохраняются и распределяются в популяции на основе закономерностей изменения частоты распространения генов. В основе таких изменений лежат закономерности наследования, открытые Г. Менделем и Т. Морганом. Знание этого позволило вывести правила распространения генов в панмиктической популяции.
Очевидно, те организмы, генотипы которых наилучшим образом обеспечивают приспособление к условиям существования, произведут большее число соответствующих гамет, чем менее приспособленные. Следовательно, частота (встречаемость) того или иного гена в популяции также будет определяться естественным отбором.
Сообщество свободноскрещивающихся генотипически различных организмов внутри вида некоторые генетики называют менделевской популяцией. Мы предпочитаем называть ее панмиктической, поскольку ее существование определяется не только законами Менделя, но и взаимодействием всех факторов эволюции, обеспечивающих свободу скрещивания организмов внутри популяции. Разнообразие генотипов панмиктической популяции — это результат мутационной и комбинативной изменчивости. Вновь возникшая мутация, чтобы стать достоянием популяции, должна сохраниться и размножиться, т. е. оказаться в составе генотипов целого ряда организмов. Любая мутация в популяции имеет свою судьбу.
Вследствие распространения большого количества разнообразных мутаций в популяции генотипы организмов оказываются насыщенными различными мутациями, которые чаще всего находятся в гетерозиготном состоянии. Так, например, что число растений, гетерозиготных по некоторым мутациям, может составлять в популяции довольно высокий процент. По мере повышения концентрации мутаций в популяции они переходят в гомозиготное состояние.
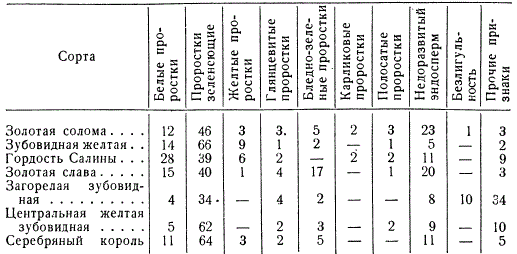
Количество растений, гетерозиготных по различным рецессивным мутациям, установленным для некоторых сортов кукурузы (в %)
Огромная насыщенность популяции мутантными генами характерна не только для культурных растений и домашних животных, но, как впервые показал С. С. Четвериков, и для природных популяций. При этом в популяции встречаются мутации, различающиеся как по своей генетической природе (генные мутации и хромосомные перестройки), так и по своему фенотипическому проявлению.
Чтобы иллюстрировать расчленение панмиктической популяции под действием отбора, рассмотрим модельный опыт с искусственно созданной гибридной популяцией, поставленный американскими генетиками Д. Джонсом и Е. Истом. Эти два исследователя скрещивали две разновидности табака, различавшиеся по длине венчика (короткий и длинный). Растения первого поколения были скрещены между собой, а из второго поколения были взяты две линии А и В со сходной изменчивостью по этому признаку.
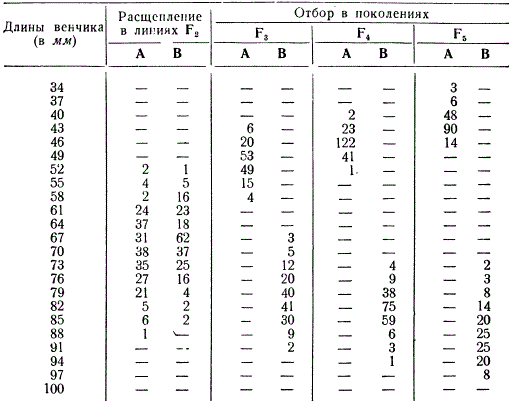
Результаты отбора (частота встречаемости) по длине венчика у табака
Длина венчика определяется многими генами, и поэтому в F2 она колебалась в этих линиях в пределах от 52 до 88 мм, В дальнейшем в потомстве взятых линий был произведен отбор в течение трех поколений: в линии А — на короткий венчик, а в линии В — на длинный венчик. В каждом поколении внутри обеих линий скрещивали отобранные формы: в линии А — с коротким венчиком, а в линии В — с длинным венчиком. Как мы видим, уже в пятом поколении линии А и В настолько сильно различались, что между ними не было никакого захождения (трансгрессии), т. е. максимальная длина венчика в линии А была меньше минимальной длины в линии В.
Следовательно, путем отбора и скрещивания отбираемых форм удается создать линии с иным выражением признака, чем у исходной популяции: отбор расчленяет популяцию на различные генотипы. В данном опыте искусственный отбор был проведен по одному признаку с преднамеренным скрещиванием растений. В природе же естественный отбор осуществляется по многим признакам и либо сохраняет и поддерживает популяцию в целостном состоянии, либо разлагает ее соответственно конкретным условиям существования.
Изучение генетики популяций ведется разными методами, основными из которых являются – цитогенетический, эколого-физиологический, математический.
Первые два метода применяются при анализе наследования в популяции — для оценки концентрации мутаций и частот, мутирования. Эколого-физиологический метод оказывается необходимым для оценки действия абиотических и биотических факторов при определении приспособительной ценности фенотипов, относящихся к генетически различным классам особей. При этом большие экспериментальные возможности открывает моделирование действия отбора в искусственно создаваемых синтетических популяциях с заранее заданными генетическими параметрами — введение в популяцию тех или иных мутаций, инверсий, транслокаций и т. д.
Математический метод позволяет давать строгое количественное описание биологических процессов. Использование электронно-вычислительных машин явилось особенно перспективным для моделирования динамики генетической структуры популяции с учетом сложного взаимодействия многих факторов. Описываемые при этом ситуации в значительной мере приближаются к отражению истинной, сложной и противоречивой картины эволюционных процессов, протекающих в естественных популяциях растений и животных.