У одноклеточных растительных организмов поглощение воды и минеральных веществ из окружающей среды, а также синтез органических соединений осуществляются настолько близко от мест их потребления, что передвижение различных веществ в пределах клетки не представляет больших затруднений.
Совершенно иное наблюдается у многоклеточных растений, в процессе эволюции которых происходила дифференциация клеток и образование отдельных тканей и органов, постепенно приспосабливавшихся к выполнению определенных функций. У покрытосеменных расчленение организма на органы достигает наибольшей степени дифференциации и специализации. В большинстве случаев их отдельные органы располагаются в физически неодинаковых условиях и выполняют совершенно различные функции. Листья осуществляют фотосинтез, для чего им необходимы минеральные вещества и вода, поступающие в растение благодаря деятельности корневой системы. В то же время продукты фотосинтеза расходуются в корнях на процессы их роста, дыхания, поглощения веществ из почвы, а также откладываются в запас. Ткани, потребляющие питательные вещества и воду, очень часто значительно удалены от тканей, их добывающих или синтезирующих. В связи с этим внутри тела растительного организма происходит передвижение различных веществ по специальной проводящей системе, которая выработалась в процессе эволюции. На очень большие расстояния (50—70 м и даже больше) передвигаются минеральные вещества, вода и ассимиляты у древесных растений.
Пути и направления передвижения веществ по растению
По каким же проводящим путям транспортируются различные вещества у высших растений? Выяснить это помогли опыты по кольцеванию растений, особенно деревьев.
Кольцевание заключается в следующем. На стволе дерева делаются два параллельных горизонтальных надреза по всей его окружности. Кора, расположенная между этими надрезами, полностью удаляется в виде кольца вплоть до древесины. Листья на окольцованном дереве продолжают фотосинтезировать и не завядают в течение длительного времени. Значит, удаление коры не нарушает нормального обеспечения листьев водой и минеральными веществами, которые поднимаются от корней к кроне не по коре, а по древесине, или по ксилеме (по ее проводящим элементам — сосудам и трахеидам). Общее направление этого тока снизу вверх, поэтому он назван восходящим. С другой стороны, опыты с кольцеванием показали, что передвижение органических веществ происходит по коре, удаление которой задерживает отток ассимилятов. Об этом свидетельствует резкое повышение содержания органических соединений в стволе выше кольца и в листьях. За счет большого количества питательных веществ усиленно разрастаются ткани, находящиеся выше кольца, в результате чего над кольцом образуется наплыв. Напротив, рост корней и участков ствола ниже кольца вскоре приостанавливается, может наблюдаться даже отмирание корней, так как отток ассимилятов из листьев вниз прекращается после удаления коры. Все это доказывает, что органические соединения передвигаются по коре, или флоэме (по ее проводящим элементам — ситовидным трубкам с клетками-спутниками). Основная масса продуктов фотосинтеза оттекает из листьев вниз к корням, поэтому этот ток и назван нисходящим.
Таким образом, с помощью кольцевания уже давно было установлено, что в растениях существуют два противоположно направленных тока веществ. Но понятия «восходящий ток» и «нисходящий ток» весьма условны. Хорошо это можно объяснить на примере нисходящего тока. Известно, что из листьев ассимиляты могут перемещаться не только вниз, к корням, но и вверх, к растущим верхушкам стебля, распускающимся цветкам и созревающим плодам. В связи с этим некоторые исследователи предлагают называть нисходящий ток просто током пластических веществ, не указывая названием на его направление. Современные экспериментальные данные, полученные с помощью радиоактивных изотопов, убедительно показывают, что термины «нисходящий ток» и «восходящий ток» условны не только в отношении направления движения, но и в том смысле, что нисходящий ток не всегда несет только органические соединения, а восходящий — только воду и минеральные соли. В частности, по ситовидным трубкам в различных направлениях перемещаются не только ассимиляты, но и неорганические вещества, а по сосудам древесины вместе с водой и элементами минерального питания могут подниматься и органические соединения.
Пути и направления движения веществ у многолетних растений существенно изменяются в зависимости от времен года. У древесных пород весной, когда они после зимнего покоя трогаются в рост, происходит интенсивная транспортировка в надземные органы органических веществ, которые были отложены в качестве запаса в корнях и стеблях. Во время весеннего сокодвижения запасные вещества быстро растворяются и в большом количестве поднимаются вверх к распускающимся почкам по ситовидным трубкам коры, а также по сосудам древесины вместе с восходящим током воды и минеральных солей. В остальное время года передвижение органических соединений осуществляется главным образом по коре, в сосудах древесины они присутствуют лишь в незначительных количествах. По мере роста листьев и образования в них ассимилятов ситовидные трубки все больше и больше становятся нисходящими путями. В течение всего лета часть продуктов фотосинтеза оттекает вниз, к корням, а другая их часть, наоборот, поднимается вверх, к точкам роста, цветкам и плодам.
С помощью меченых атомов обнаружено, что направление оттока ассимилятов в большой мере зависит от местоположения листьев, которые по-разному снабжают питательными веществами отдельные органы. Нижние листья до самого пожелтения обеспечивают ассимилятами корневую систему, средние — развивающиеся плоды и пазушные побеги, а верхние — растущую верхушку главного побега. Таким образом, транспорт органических веществ по флоэме носит местный характер, т. е. от каждого листа продукты фотосинтеза поступают преимущественно в определенный участок растения.
Основные транспортируемые вещества
Большая часть передвигающихся ассимилятов приходится на углеводы, содержание которых во флоэмном соке может достигать 90%, остальное составляют различные аминокислоты, амиды, спирты, органические кислоты и некоторые другие вещества. По современным представлениям, главным транспортным соединением у большинства растений является сахароза. Наряду с сахарозой почти всегда передвигаются и ее производные — олигосахариды, такие, как рафиноза, стахиоза и вербаскоза. Эти вещества образуются в результате присоединения к сахарозе соответственно одного, двух или трех остатков галактозы, как это показано на рисунке. У некоторых древесных пород, например из семейства липовых, вязовых, самшитовых и некоторых других, указанные олигосахариды преобладают даже над сахарозой по содержанию во флоэмном соке.
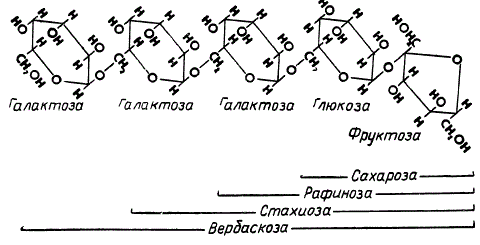
Олигосахариды, образующиеся путем присоединения к сахарозе одного, двух или трех остатков галактозы
Помимо сахарозы и олигосахаридов, транспортируемыми веществами служат спирты, имеющие в цепочке по шесть атомов углерода: маннит, сорбит и дульцит. У растений из семейств сиреневых и ясеневых обычно содержится маннит, у представителей яблоневых и сливовых — сорбит, а у бересклетовых — дульцит. В то же время во флоэмном соке растений различных видов свободные гексозы практически отсутствуют. Все это позволило акад. А. Л. Курсанову предположить, что углеводы, передвигающиеся на большие расстояния по растению, не должны принимать непосредственного участия в обмене веществ проводящих элементов коры. Преимущество сахарозы и олигосахаридов в качестве транспортных веществ как раз и заключается в том, что входящие в их состав гексозы находятся в связанном состоянии и поэтому защищены от использования в обмене веществ ситовидных трубок. То же самое можно сказать и о вышеупомянутых шестиатомных спиртах, которые образуются путем восстановления гексоз. Таким образом, гексозы на время транспортировки по растению преобразуются в нередуцирующие сахара (сахарозу и олигосахариды) или в спирты, т. е. в такие формы, которые не используются на дыхание и другие превращения в ситовидных трубках.
Интересно отметить, что сами ситовидные трубки имеют своеобразные приспособления для транспортировки углеводов. Одно из таких приспособлений состоит в уменьшении количества митохондрий ниже нормы, благодаря чему в них снижается углеводный обмен. Другая морфологическая особенность строения ситовидных трубок, обеспечивающая транспортировку азотистых веществ на далекие расстояния, заключается в следующем. Уже на ранних этапах развития их клетки утрачивают ядро и ядрышко. Такое изменение, по-видимому, приспособительно: оно ограничивает нуклеиновый обмен в ситовидных клетках, а следовательно, снижает расход аминокислот на биосинтез белка, в результате чего большая часть аминокислот передвигается по растению.
Использованию ассимилятов, как правило, предшествует их отток. Непосредственно на месте синтеза — в листьях расходуется лишь очень незначительная часть образовавшихся здесь органических веществ, основная же масса их поступает в проводящую систему и транспортируется в различные органы растения. Отток ассимилятов начинается с перехода их из фотосинтезирующих клеток в окончания проводящих путей.
А. Л. Курсановым с сотрудниками установлено, что выход углеводов, аминокислот и некоторых других веществ из фотосинтезирующих клеток в проводящую систему осуществляется избирательно. Это, следовательно, не результат пассивной диффузии, а идущий с затратой энергии активный процесс, в котором большую роль играет обмен веществ. Избирательность означает, что переход различных веществ в проводящую систему происходит в неодинаковой степени и с различной скоростью, поэтому одних веществ поступает во флоэму больше, других — меньше, а некоторые соединения практически вообще не выходят из клеток листа. Выше уже говорилось о том, что углеводы транспортируются главным образом в виде сахарозы и олигосахаридов, а не в виде гексоз. Причем сахарозы во флоэмном соке содержится значительно больше, иногда в 5—6 раз, чем в тканях листа, что. также свидетельствует об избирательном характере поступления углеводов в ситовидные трубки. Еще более наглядным примером, иллюстрирующим избирательный выход веществ во флоэму, может служить отток и транспортировка аминокислот. По данным А. Л. Курсанова, у ревеня в середине лета особенно активно транспортируется треонин, который легче других аминокислот поступает в проводящую систему. На его долю во флоэмном соке приходится в этот период около 25—30% от общего количества аминокислот, в то время как в самих фотосинтезирующих клетках содержание треонина очень низкое — лишь около 1—3%. Аспарагиновая же кислота почти совсем не переходит в окончания проводящей системы, хотя содержание ее в фотосинтезирующих тканях листа намного превышает количество треонина в них.
Скорость оттока и передвижения ассимилятов
Сравнительно недавно с помощью радиоактивных изотопов удалось определить, что отток веществ из зеленых клеток листа в окончания проводящих пучков осуществляется с большой скоростью, несмотря на то что движение происходит не по специализированным клеткам листовой паренхимы. На растениях ячменя и сахарной свеклы было показано, что после кратковременного пребывания листьев в атмосфере с добавлением радиоактивной углекислоты (С14O2) «меченые» продукты фотосинтеза перемещаются из мест их образования — зеленых клеток листа в проводящие пучки флоэмы всего лишь за несколько минут. Еще более быстро ассимиляты транспортируются по специально приспособленным для этой цели ситовидным трубкам. «Меченые» атомы позволяют сравнительно легко обнаруживать продукты фотосинтеза в различных частях растения и достаточно точно определять скорость их передвижения. Этим способом было установлено, что как у травянистых, так и у древесных растений скорость транспортировки ассимилятов по флоэме в среднем колеблется от 80 до 130 см/ч, т. е. она в тысячи раз превосходит скорость диффузии. Сравнительно недавно удалось обнаружить, что небольшие количества органических веществ могут перемещаться по растению чрезвычайно быстро, со скоростью 50—70 м/ч. Однако эти интересные данные, полученные пока что для сои и картофеля, нуждаются в дальнейшей экспериментальной проверке на растениях разных видов.
Распределение ассимилятов и влияние различных условий на их отток
Рост и развитие отдельных органов растения в большой степени зависит от количества и вида поступающих к ним питательных веществ. Об этом свидетельствуют хотя бы такие хорошо известные факты, как неравномерный рост аналогичных органов (например, плодов), расположенных вблизи друг от друга, замедление или даже полная приостановка роста одних органов в результате усиления роста других и т. п. Питательные вещества никогда не распределяются равномерно по всему растению, так как на каждой фазе развития ток ассимилятов направляется преимущественно к активным центрам их потребления. Такими центрами могут быть наиболее интенсивно растущие или запасающие ткани и органы, а также органы с повышенным обменом веществ, усиленно расходующие органические вещества. На протяжении жизни растения активными центрами потребления питательных веществ становятся то одни, то другие органы, благодаря чему направление и скорость оттока ассимилятов из листьев, а следовательно, и распределение их по растению значительно изменяются. У злаков в фазе кущения, например, активными центрами потребления питательных веществ среди надземных органов являются листья, так как именно они интенсивно растут в этот период; в фазе выхода в трубку активными центрами становятся стебли и соцветия, а после цветения наибольший приток органических веществ наблюдается к новым центрам — к созревающим плодам.
Таким образом, отток ассимилятов и их передвижение регулируются самим растением при непосредственном участии проводящей системы, которая обеспечивает связь отдельных органов друг с другом. По современным представлениям, благодаря постоянному току веществ растительный организм функционирует как единое целое, в котором налицо физиологическая взаимосвязь всех его частей. Проводящие ткани выполняют роль регуляторной системы, осуществляющей координацию и управление на расстоянии деятельностью различных органов. Большую роль при этом играют, по-видимому, электроимпульсы, с помощью которых у растений, как и у животных, происходит передача информации на расстояние при изменении условий внешней среды.
И. И. Гунаром, А. М. Синюхиным и некоторыми другими исследователями было установлено, что электроимпульсы возникают при воздействии различных факторов внешней среды на корневую систему, листья или стебли. Затем они распространяются вдоль стебля преимущественно по проводящим пучкам — с довольно большой скоростью (10—80 см/мин) и таким образом передаются на другие органы. Электроимпульсы, возникшие в корнях, могут передаваться на листья, вызывая ту или иную реакцию в ответ на доставленную из корней информацию. В частности, удалось выяснить, что электросигналы, возникшие в корнях при воздействии на них ионов калия, распространяются на листья, в результате чего значительно усиливается отток продуктов фотосинтеза к корневой системе. В ответ на информацию, поступившую из одних органов в другие, у последних может изменяться интенсивность физиологических процессов, причем воздействие одних факторов обусловливает усиление, а других, наоборот, понижение интенсивности. Так, при действии на корни индолилуксусной кислоты, хлористого калия или других питательных солей наблюдается усиление фотосинтеза, тогда как под влиянием искусственного стимулятора роста 2,4 ДУ или тепловой обработки интенсивность этого процесса снижается.
Большое значение для оттока и передвижения продуктов фотосинтеза имеют ауксины, гетероауксин, гиббереллины и другие гормоны растений, называемые фитогормонами (от греч. фитон — растение). Определение содержания веществ типа ауксина, способных стимулировать рост, показало, что ток ассимилятов направляется преимущественно к тканям, наиболее богатым этими веществами. В каждом растущем или запасающем органе накапливаются фитогормоны, под влиянием которых значительно усиливается поступление органических веществ и тем самым регулируется направление и скорость их оттока, а в конечном счете и распределение между различными частями растения. Это показывает, что человек располагает огромными возможностями для искусственного управления процессами оттока и передвижения питательных веществ с целью повышения урожая сельскохозяйственных культур. Изменяя количество фитогормонов в тех или иных органах, можно не только усилить или ослабить отток, но и существенным образом повлиять на направление транспортировки ассимилятов.
Искусственное обогащение гетероауксином нижней части стебля пшеницы, как показала Н. И. Якушкина, приводит к тому, что сахара передвигаются в направлении, противоположном обычному. Если при нормальных условиях ассимиляты поднимаются главным образом к верхней части стебля и к колосу в полном соответствии с наибольшим содержанием в них веществ типа ауксина, то у обработанных растений колосья не только перестают быть активными центрами потребления питательных веществ, но из них даже осуществляется отток углеводов к нижним частям стебля, и теперь уже основная масса сахаров передвигается в нисходящем направлении.
Перераспределение выработанных растением органических веществ и усиление роста одних органов за счет других может также достигаться с помощью искусственных стимуляторов роста. Уже через 5 дней после опрыскивания цветков томатов искусственным стимулятором роста 2,4 ДУ приток сахаров к ним увеличивается в 5 раз, а азотистых веществ — более чем в 2 раза по сравнению с контрольными растениями (не опрыскиваемыми стимулятором). Причем особенно сильно возрастает приток ассимилятов к завязям из листьев нижнего яруса, которые в обычных условиях обеспечивают продуктами фотосинтеза главным образом корневую систему и практически не участвуют в снабжении цветков и плодов. Кроме того, после опрыскивания растений стимуляторами роста может расширяться круг листьев, снабжающих плоды, что обеспечивает более продуктивное использование ассимилятов для формирования урожая.
Уже накоплено немало данных, свидетельствующих о том, что скорость и направление движения продуктов фотосинтеза по растению могут существенно изменяться под влиянием различных факторов внешней среды. В этой связи достаточно вспомнить хотя бы о значении света для процесса клубнеобразования у картофеля. Хорошо известно, что при искусственном затемнении верхней части побега картофеля ток пластических веществ становится прямо противоположным. Основная масса ассимилятов направляется из листьев не вниз, как обычно, а, наоборот, в верхнюю часть побега, где начинается усиленное клубнеобразование, при этом в подземных частях растения формирование клубней значительно замедляется из-за недостатка пищи.
Обычно скорость передвижения веществ по флоэме возрастает с повышением температуры примерно до 20—30° С; при более высокой температуре этот процесс весьма замедляется или даже полностью прекращается, так же как и при сильном охлаждении.
С помощью радиоактивных изотопов было установлено, что хорошие условия водоснабжения содействуют оттоку и транспортировке органических веществ, а продолжительная и сильная засуха оказывает противоположное действие. Однако непродолжительная или неглубокая засуха может даже стимулировать отток сахаров и азотистых веществ в зависимости от степени обезвоживания.
Одним из главных условий передвижения ассимилятов, помимо нормального водообеспечения растений, является наличие достаточного количества кислорода, необходимого для дыхания клеток ситовидных трубок.
Изучение влияния элементов минерального питания показало, что при соприкосновении корневой системы с растворами минеральных солей усиливается отток продуктов фотосинтеза из листьев в сторону корней, а также ускоряются процессы переработки поступивших в них веществ. В отсутствие или при недостатке элементов минерального питания не только ухудшается снабжение корней органическими веществами, но и замедляются процессы превращения этих веществ. Очень большое значение для передвижения ассимилятов имеют как макро-, так и микроэлементы. Под влиянием бора, например, а особенно комбинации бора с марганцем, резко возрастает скорость оттока сахаров и улучшается снабжение репродуктивных органов пластическими веществами.
Одним из основных процессов, определяющих величину и качество урожая, является приток ассимилятов к хозяйственно ценным органам. В связи с этим первостепенное значение приобретает разработка действенных методов управления оттоком и перераспределением питательных веществ между отдельными частями растения. Заманчивые перспективы в этом отношении открываются благодаря глубокому изучению закономерностей передвижения продуктов фотосинтеза при различных условиях внешней среды. Уже известно, что посредством изменения водного или температурного режима, условий освещения, минерального питания, а также посредством чеканки, подрезки, прищипывания побегов и других агротехнических приемов можно регулировать распределение питательных веществ в пределах растительного организма. Большую ценность с практической точки зрения представляют способы обработки растений различными химическими препаратами, в первую очередь искусственными стимуляторами роста, под влиянием которых значительно изменяется не только скорость и направление передвижения, о чем мы уже говорили, но и состав транспортируемых ассимилятов. Весьма эффективной для «перекачки» органических веществ из листьев в другие органы с целью увеличения их урожая может оказаться и обработка дефолиантами, т. е. веществами, ускоряющими старение и опадение листьев.
У многих сельскохозяйственных культур урожай зерна, клубней, луковиц и т. д. убирают до отмирания листьев, и большие количества содержащихся в них различных органических веществ остаются обычно неиспользованными. Под влиянием дефолиантов в обработанных растениях происходят физиологические изменения, аналогичные тем, которые характерны для старения листьев перед естественным листопадом: усиливаются процессы распада белков, крахмала и других сложных органических соединений. Образующиеся при этом аминокислоты, сахара и т. д. оттекают из постепенно отмирающих листьев к плодам, корнеплодам или другим запасающим органам, в результате чего урожай их увеличивается.
Таким образом, обработка растений дефолиантами, как и стимуляторами роста, может иметь немаловажное хозяйственное значение. И несомненно, что дальнейшее изучение проблемы оттока ассимилятов позволит в недалеком будущем реализовать большие перспективы, которые определились в отношении активного управления формированием урожая сельскохозяйственных культур с помощью применения химических препаратов и других агротехнических мероприятий.
Источник: Н.Н. Овчинников, Н.М. Шиханова. Фотосинтез. Пособие для учителей. Изд-во «Просвещение». Москва. 1972