С обогащением атмосферы свободным кислородом благодаря все возрастающей деятельности фотосинтетиков создается озоновый экран Земли (когда уровень O2 достигает примерно 1% от современного состояния). До. возникновения озонового экрана из-за губительного действия УФ-лучей отбор даже среди первичных многоклеточных растений, в том числе и на суше, шел в сторону безлистности.
Важным следствием насыщения атмосферы O2 было чрезвычайное повышение энергетических процессов живых организмов путем метаболического использования свободного кислорода. Это событие имело грандиозное значение для прогрессивного развития живой природы и сопровождалось усилением связи процессов фотосинтеза и дыхания. Таким путем создалась возможность усиления круговорота энергии на Земле, энергетического обеспечения хлоропластов деятельностью митохондрий. И путь совершенствования живых организмов в этом направлении был длительным и ступенчатым.
Большая часть современных живых организмов не может обойтись без кислорода, используемого для дыхания. Напротив, на начальных этапах развития жизни на Земле кислород выступал как опасный для нее фактор. Это становится понятным, если принять во внимание его способность активно поддерживать горение и вызывать окисление разных веществ. Кислород в природе образуется при разложении воды, как и в случае фотосинтеза (фотолиз воды). Исторически сложилось так, что организмы, выделяющие кислород, развиваются раньше, чем потребляющие его. Например, многие даже современные цианеи в процессе фотосинтеза выделяют O2, хотя у них нет механизмов аэробного обмена.
Существует мнение о самопроизвольном возникновении биоэнергетических механизмов обмена (R. F. Fox, 1982). В соответствии с этим энергообеспечение первых живых существ есть не что иное, как использование механизмов, сложившихся в неживой природе. При этом неорганический пирофосфат рассматривается как древнейшая «конвертируемая» валюта. Возникновение протонного потенциала связывают с реакцией оксиредукции между НАД ∙ Н и Fe2+ на мембранах. Роль трансмембранного переносчика в последней реакции отводится убихинонам.
Как известно, ныне живущие бактерии отличаются огромным разнообразием биоэнергетических процессов, у них значительно варьирует ферментативный аппарат и в дыхании участвует несколько терминальных оксидаз (Э. Брода, 1978).
Остановимся на начальных этапах извлечения энергии живыми существами первичной Земли. Естественно, что об этом можно судить только по косвенным подходам — сравнению процессов энергообмена у современных наиболее примитивных существ, являющихся персистированными представителями древних форм жизни.
Многие данные не оставляют сомнения в том, что наиболее древние формы жизни нашей планеты (протобионты) были гетеротрофами, извлекали энергию из готовых органических веществ в результате субстратного фосфорилирования (гликолиза). Однако некоторые авторы не считают процессы гликолиза самыми примитивными проявлениями широко распространенного анаэробного энергообмена — брожения. Так, брожению могли предшествовать довольно простые реакции энергетического обмена наподобие обмена клостридий, которые для синтеза АТФ используют ограниченное число реакций с участием НАД, КоА, НАДФ и ферредоксина. Наиболее примитивные организмы в ходе субстратного фосфорилирования перерабатывают органические соединении без предварительной их активации с помощью АТФ, так как субстраты обладают сильно выраженными восстановительными свойствами (Э. Брода, 1978; М. В. Гусев, А. А. Минеева, 1985; Т. В. Чиркова, Г. Ж. Валиханова, 1985).
Дело в том, что на ранних этапах развития жизни, когда в среде изобиловали различные органические вещества абиогенного происхождения, было найдено простое решение для поддержания и воспроизведения жизни — извлечение энергии из готовых органических веществ. Смысл этого можно понять, если принять во внимание отсутствие у протобионтов развитых собственных систем для получения энергии. Словом, организмы не располагали возможностью извлекать энергию из собственных химических реакций ввиду их чрезвычайной упрощенности и примитивности своей организации. Такой способ получения энергии предполагает наличие механизма гомоферментативных реакций по типу молочнокислого брожения (ключевой фермент — фосфофруктокиназа), где гликолитический распад сахаров осуществляется (у бактерий в цитоплазме) вне мембранных структур. Напомним, что при этом высвобождение энергии из субстрата и фиксация ее в виде АТФ осуществляются метаболической перестройкой органических молекул, в состав которых входили не полностью окисленные (или восстановленные) атомы углерода. Энергии при этом выделяется мало ввиду образования в Качестве конечных продуктов сильно восстановленных соединений. Поэтому синтезируются лишь две молекулы АТФ на молекулу гексозы.
Ввиду опасности быстрого исчерпания субстрата для фосфорилирования на ранних этапах развития жизни отбор организмов среди протобионтов пошел по пути опробования разных субстратов в энергетических целях. По принципу Гаузе эволюция пошла в направлении использования разных субстратов как источников энергии, что способствовало повышению емкости первичных водных биоценозов и надежности воспроизведения организмов. Адаптивная радиация гетеротрофных первичных организмов сопровождалась специализацией их по переработке разных субстратов.
Нет необходимости описывать известные реакции субстратного фосфорилирования. Однако заметим, что наиболее важной из них является окисление 3ФГК: здесь клетка как бы «возвращает» две молекулы АТФ, ранее ею же затраченные на активирование субстрата. Эта реакция протекает вне определенных структур клетки, и запасание Выделяемой энергии (энергетическая эффективность) в виде АТФ составляет 40%. Другим свидетельством примитивности субстратного фосфорилирования служит недифференцированность донора и акцептора электронов. При этом субстрат выступает в роли как донора, так и акцептора электронов, что ограничивает его окислительные возможности.
Энергетические ограничения гликолиза связаны и с постепенным ингибированием активности его ключевого фермента по мере накопления АТФ, цитрата и Н+. Торможение конечных реакций обусловлено при брожении отсутствием регенерации НАД+, что приводит к застопориванию превращения триозофосфата и образования АТФ (Т. В. Чиркова, Г. Ж. Валиханова, 1985). В то же время субстратное фосфорилирование ввиду простоты механизма извлечения энергии, относительно высокой энергетической эффективности (даже при низком выходе энергии) и разнообразия самих способов превращения субстратов нашло широкое использование в последующей эволюции жизни на Земле. Оно стало основой для развития более совершенных форм метаболизма. Исторически наиболее древней следует признать функцию АТФ как источника создания электрохимического потенциала на мембране ΔμН+. Позднее возникла возможность синтеза АТФ за счет ΔμН+ путем изменения направления работы протонного АТФ-азного комплекса: АТФ ↔ ΔμН+.
В эволюционном плане предшественниками АТФ служат пирофосфаты как источники биохимических реакций. Так, на анаэробе Desulfomaculatus nigrificans показана возможность роста в присутствии пирофосфата:
Ацетат + пирофосфат + АДФ → Ацетат + ортофосфат + АТФ
Как известно, в морской воде отсутствует пирофосфат и его функцию там выполняет полифосфат. Многие анаэробные формы с древнейших времен продолжают использовать пирофосфаты как, источник энергии (F. Lipmann, 1984). В этом смысле преимущественное использование АТФ ↔ АДФ (а не АДФ ↔ АМФ, в термодинамическом отношении также выгодные) в энергетических целях вызывает удивление. «Выбор» в ходе эволюции АТФ ↔ АДФ объясняют преимущественным участием в реакциях их хелатных соединений с магнием, облегчающих взаимодействие с ферментами (J. Е. Wilson, 1984). Возможно, что при этом сыграло роль и химическое предрасположение — относительная легкость синтеза адениновых нуклеотидов на стадии биопоэза.
Сказанное о роли субстратного фосфорилирования и механизмов энергообмена у древних форм подтверждает мнение Ч. Дарвина о том, что «при очень простых жизненных условиях высокая организация не оказала бы никакой услуги…». Действительно, условия первичных биоценозов Земли были крайне упрощены. Им как раз соответствовали примитивная организация протобионтов и система их энергообеспечения. В этом проявляется целесообразность организации протобионтов, являющаяся результатом действия естественного отбора.
На подобной основе, вероятно, и появляется гликолиз, затем возникает механизм мембранного фосфорилирования, представляющий собой важный этап биоэволюции. Мембранное фосфорилирование в редокс-цепи связано со строгой пространственной организацией его звеньев. Впервые оно складывается у фототрофных прокариот. Поэтому фототрофия эволюционно предшествовала аэробиозу. У древних бактерий функционирует глиоксилатный путь, представляющий собой сокращенный вариант цикла трикарбоновых кислот (ЦТК). ЦТК также относится к древнему приобретению жизни, который до момента появления кислорода в атмосфере не нашел эффективного применения (М. В. Гусев, Г. Б. Гохлернер, 1980). О первичности анаэробного обмена свидетельствуют многообразные метаболические превращения, даже у высших организмов совершающиеся в отсутствие O2.
На следующем этапе эволюции возникает ПФП метаболизма, играющий важную роль в образовании нуклеиновых кислот, метаболически активного водорода в форме НАДФ ∙ Н и предшественников ряда аминокислот. Впервые ПФП появляется также у фототрофов. У сбраживающих бактерий ПФП не обнаружен, у других бактерий он сформировался позже фотосинтеза.
Значение ПФП велико прежде всего в образовании активированного водорода путем расщепления воды и использовании его для восстановления НАДФ, образовании пентоз, необходимых для синтеза нуклеиновых кислот. В эволюционном плане эти события способствовали усилению независимости организмов от внешней среды, что рассматривается как один из основных результатов прогресса живой природы (И. И. Шмальгаузен). Преимущества ПФП обусловлены возможностью циклического его осуществления, где из шести молекул глюкозо-6-фосфата только одна расщепляется до CO2.
В то же время ПФП обладает рядом примитивных особенностей (встречается у анаэробов), его ферменты (транскетолаза и трансальдолаза) распространены у всех живых организмов и включают некоторые реакции брожения. Не исключено, что ПФП мог сложиться тогда, когда организмы перешли к самостоятельному синтезу гексоз, т. е. параллельно с формированием фототрофии (М. В. Гусев, Г. Б. Гохлернер, 1981). Несмотря на заметную метаболическую гибкость ПФП, он не дает больших преимуществ в синтезе АТФ (А. Ленинджер, 1974), так как у эукариот в митохондриях НАДФ не включается в дыхательную цепь и не используется для синтеза АТФ. Однако НАДФ ∙ Н принимает участие в трансгидрогеназной реакции и в генерации мембранного потенциала:
НАДФ ∙ Н + НАД+ → НАДФ+ + НАД ∙ Н,
что по своему направлению сходно с результатами аэробного дыхания и гидролиза (В. П. Скулачев, 1974).
У цианей ЦТК выражен неполно: преобладает ПФП, а водород вовлекается в цепь в виде НАДФ ∙ Н. Точно так же некоторые аэробные бактерии (уксуснокислые) осуществляют неполное окисление органических веществ. Водород, высвобождаемый в результате дегидрирования субстрата, вовлекаясь в процессы окислительного фосфорилирования, превращается в воду. Полный ЦТК впервые формируется у фототрофных анаэробных бактерий.
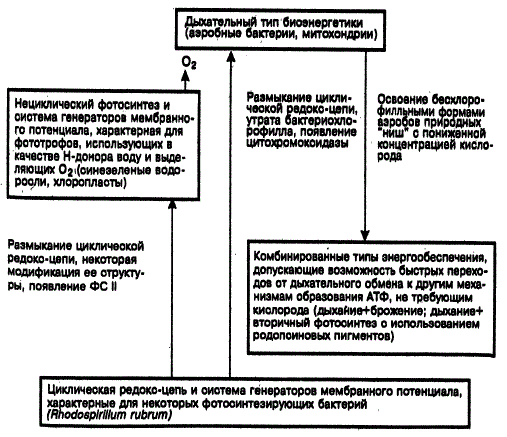
По указанным причинам у современных бактерий можно встретить разнообразные сочетания процессов фотосинтеза, аэробного дыхания и гликолиза. Переход анаэробных бактерий к аэробному обмену (т. е. к дыхательной цепи) был связан не столько с изменением структуры мембран, сколько с усилением прочности связи ферментов с ними. Это показано на примере сравнительного изучения локализации ряда дегидрогеназ у современных аэробных и анаэробных бактерий. Дыхательная цепь впервые появляется у прокариот и исторически складывается путем замены системы бактериохлорофилл первичный акцептор цитохромами a и a3, что привело к использованию O2 как конечного акцептора электронов.
М. В. Гусев и Г. Б. Гохлернер (1980) так представляют последовательность этапов энергетического обмена у бактерий: брожение → фотосинтез → дыхание → вторичный анаэробиоз → вторичный фотосинтез (микроаэрофилия) нитратное «дыхание» → хемолитотрофия. Микроаэрофилия и нитратное «дыхание» возникают благодаря вторичному продвижению части аэробов в бедные кислородом экологические ниши. Перечисленные этапы обмена могли возникать конвергентно и неодновременно у разных прокариот путем видоизменения реакций брожения, гликолиза и ПФП, которые и лежат в основе дыхания представителей всех последующих царств и определяют сходство равновесных реакций кинетического контроля их метаболизма.
Дыхательные ферменты растений близки к таковым животных, у которых нет кровеносной системы и постоянной температуры тела. У таких животных и растений на завершающем этапе дыхания отмечена лабильность и многокомпонентность процесса. Но растения, сами вырабатывая кислород, меньше страдают от анаэробиоза и отличаются цианидоустойчивым дыханием (ЦУД). Это дыхание является следствием включения альтернативных путей транспорта электронов с участием флавопротеинов. Кроме того, дыханию растений свойственна полифункциональность одних и тех же ферментов и связь с процессами фотосинтеза. ЦУД представляет собой компонент общего дыхания, оно повышается у растений в онтогенезе при переходе к репродуктивной фазе и в стрессовых условиях. Эволюционно возникновение ЦУД можно представить как результат отбора растений по активности цитохромоксидаз и перехода к преимущественному использованию фруктозы и гексозы в качестве субстратов дыхания в стрессовых условиях (Н. W. Janes, Chin Chin-Kok, 1981).
Для понимания особенностей дыхания разных организмов следует учитывать и результаты морфологической эволюции. Известно, что на разных уровнях организации живых существ вопрос о снабжении кислородом решался неоднозначно (способы обеспечения им зависят от толщины кутикулы и величины поверхности тела).