Половое размножение организмов связано с их морфологической и физиологической половой дифференциацией (половой диморфизм) и половым процессом.
Половой процесс характеризуется системой приспособительных механизмов:
- образованием мужских и женских гамет,
- их слиянием в процессе оплодотворения (сингамия),
- объединением ядер (кариогамия),
- синаупсисогомологивдных хромосом в мейозе и перекомбинацией наследственных факторов.
Цикл полового размножения охватывает период от момента формирования половых клеток до их нового воспроизведения в следующем поколении.
Оплодотворением принято называть побуждение яйца к развитию в результате кариогамии. Оплодотворение представляет собой процесс необратимый — оплодотворенное однажды яйцо не может быть оплодотворено вновь. Сингамия и кариогамия составляют сущность процесса оплодотворения. Однако у некоторых видов воспроизведение нового поколения осуществляется на основе только женской гаметы — яйцеклетки без оплодотворения (девственное размножение). В этом случае половое размножение также заканчивается созреванием гамет. Оба эти способа размножения могут чередоваться у одного и того же вида.
В процессе оплодотворения осуществляются следующие важные генетические явления, необходимые для существования вида:
- восстановление диплоидного набора хромосом, а в пределах диплоидного набора — парности гомологичных (материнских и отцовских) хромосом, разошедшихся в мейозе в процессе образования половых клеток у родительских организмов;
- обеспечение материальной непрерывности между следующими друг за другом поколениями;
- объединение в одном индивидууме наследственных свойств материнского и отцовского организмов.
Для обеспечения оплодотворения необходимо одновременное созревание гамет материнского и отцовского организма. У перекрестноопыляющихся растений созревание мужских и женских половых клеток может не совпадать во времени, и это несоответствие служит приспособительным механизмом, препятствующим самоопылению. Возможно, что несоответствие во времени созревания половых клеток у разных полов одного вида является одним из путей возникновения перекрестного опыления.
Оплодотворение у животных
Процесс оплодотворения у животных можно разделить на несколько фаз.
Первая фаза начинается с того, что сперматозоид либо прикрепляется к любой точке поверхности яйца, либо проникает в нее через микропиле. Момент соприкосновения головки сперматозоида с яйцом является начальным в цепи химических реакций. Эту фазу называют фазой активации яйца. В норме активацию яйца вызывают сперматозоиды своего вида. В некоторых случаях (у червя Rhabdites monohystera) сперматозоиды могут активировать яйцо, но при этом мужское ядро не сливается с материнским. Такое явление называют псевдогамным оплодотворением.
Вторая фаза процесса оплодотворения начинается после проникновения в яйцо, одного, а у некоторых животных и нескольких сперматозоидов. Проникший сперматозоид «готовится» к слиянию с женским ядром и последующему митозу: ядро сперматозоида постепенно набухает и приобретает вид интерфазного ядра. Такое ядро называют семенным, или мужским, пронуклеусом.
К моменту соприкосновения сперматозоида с яйцом и проникновения его внутрь ядро яйцеклетки у разных животных может находиться на разных стадиях деления созревания. Ядро яйцеклетки, готовое к слиянию с ядром сперматозоида, называют женским пронуклеусом. Собственно оплодотворение, т. е. слияние отцовского и материнского пронуклеусов, возможно лишь после окончания мейоза.
Проникновение сперматозоида может происходить на стадиях:
- ооцита I с покоящимся ядром
- ооцита I в стадии метафазы I
- ооцита II в стадиях мета — или анафазы II
- зрелой яйцеклетки
У иглокожих и кишечнополостных сперматозоид может проникать в яйцеклетку после завершения мейоза. Такое оплодотворение называют оплодотворением типа морского ежа. После проникновения сперматозоида в яйцо его ядро вскоре соединяется с женским ядром; ядро зиготы приступает к первому делению — дроблению яйца.
У бесчерепных (ланцетник) и всех позвоночных проникновение сперматозоида в яйцеклетку происходит, как правило, во время метафазы II. У асцидий, двустворчатых моллюсков и ряда других животных сперматозоид проникает в яйцеклетку на стадии метафазы I, а у губок, аскарид и некоторых других животных — на стадии ооцит I, т. е. до наступления мейоза. Этот тип оплодотворения называют типом аскариды. Проникший в цитоплазму яйца сперматозоид «ожидает» в стадии покоя окончания второго мейотического деления яйца.
В акте оплодотворения два гаплоидных пронуклеуса сливаются в одно ядро. Кариогамия дает начало новому качественному процессу — развитию зиготы. Этот момент является кульминационным пунктом процесса полового размножения. В результате кариогамии, гомологичные хромосомы, разошедшиеся в мейозе предыдущего поколения, вновь воссоединяются в одном ядре зиготы.
Для понимания ряда важных генетических явлений необходимо знать, какие элементы сперматозоида проникают в яйцеклетку. Раньше считалось, что цитоплазма сперматозоида и ее органоиды не попадают в яйцеклетку. В настоящее время все больше накапливается фактов в пользу того, что в цитоплазму яйцеклетки у млекопитающих проникает не только головка (ядро) сперматозоида, но и его шейка и даже хвостовая часть. Если это подтвердится, то взгляды на роль цитоплазмы мужского организма в передаче его свойств потомству должны быть пересмотрены. Впрочем, генетических данных на этот счет пока нет; известны лишь факты передачи вирусных заболеваний.
Вместе с ядром сперматозоида в цитоплазму яйцеклетки проникает центриоль, которая через некоторое время образует центросферу, дающую начало веретену дробления.
Приведенное общее описание оплодотворения у животных в деталях может варьировать у разных видов. Вследствие этих изменений процесс оплодотворения у каждого вида может протекать специфично, препятствуя межвидовому скрещиванию.
Оплодотворение у растений
У растений так же, как и у животных, сущность оплодотворения сводится к слиянию двух гаплоидных ядер.
Оплодотворение у растений в принципе сходно с таковым у животных, однако существование у растений гаметофита привело к появлению у них и некоторых особенностей.
Цитологический механизм этого процесса у голосеменных был создан русским ботаником Н. Н. Горожанкиным в 1880 г., а у покрытосеменных — Е. Страсбургером в 1884 г. Е. Страсбургер охарактеризовал оплодотворение у покрытосеменных следующим образом:
- процесс оплодотворения включает в себя слияние ядра мужской и женской гамет,
- цитоплазма гамет не имеет отношения к оплодотворению,
- ядро спермия и ядро яйцеклетки суть настоящие ядра.
Слияние спермия с ядром яйцеклетки и является собственно актом оплодотворения, в результате которого образуется зигота с диплоидным набором хромосом.
Выше было сказано, что микрогаметогенез завершается образованием двух спермиев, которые возникают или в пыльцевом зерне, или в пыльцевой трубке при прорастании пыльцевого зерна. Время начала прорастания зерен после попадания их на рыльце у разных растений варьирует в зависимости, от внешних условий и состояния рыльца и пестика. Так, например, у свеклы прорастание пыльцевых зерен начинается через 2 ч, у кок-сагыза — через 5 мин, а у кукурузы, сорго и других растений происходит почти немедленно.
Первым признаком прорастания пыльцевого зерна является увеличение его объема. Обычно из одного пыльцевого зерна образуется одна трубка, но у некоторых растений (мальвовые, тыквенные) из одного зерна образуется несколько трубок, однако полного развития достигает лишь одна из них. Характер роста пыльцевых трубок определяется наследственными свойствами растений. К. Корренсом у MeiaridrTum (дрёма) было обнаружено, что при одновременном прорастании на рыльце нескольких пыльцевых зерен скорость роста пыльцевых трубок нередко зависит от их числа: чем больше их, тем медленнее они прорастают, при этом наблюдается конкуренция.
Пыльцевая трубка, дорастая до микропиле, приходит в соприкосновение с той частью зародышевого мешка, где находится яйцевой аппарат — яйцеклетка и синергиды. Впрочем, у некоторых растений пыльцевая трубка подходит к зародышевому мешку через халазальную часть семяпочки.
Передвигающиеся по пыльцевой трубке по мере ее роста два генеративных ядра — спермия после разрыва трубки вместе с ее содержимым попадают внутрь зародышевого мешка. Спермин могут быть округлой, штопорообразной формы, иногда разрыхленные, с видимыми хромосомными нитями и др. Ядра их в этот момент, как правило, находятся в стадии телофазы. Из двух проникших в зародышевый мешок спермиев один спермий внедряется яйцеклетку и сливается с гаплоидным ядром последней. Слияние ядра спермия с ядром яйцеклетки является центральным моментом оплодотворения у растений.
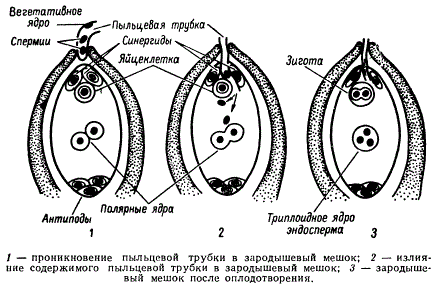
Схема двойного оплодотворения у растений
У растений так же, как и у животных, готовность к слиянию мужского и женского ядер может быть различной. Условно можно считать, что у растений имеются два типа оплодотворения: тип сложноцветных, аналогичный типу морского ежа у животных, и тип лилейных, аналогичный типу аскариды. В первом случае (тип сложноцветных) ядро спермия проникает в зрелую яйцеклетку в состоянии незавершенной телофазы, растворяет оболочку ядра яйцеклетки и переходит в интерфазное состояние. Во втором случае (тип лилейных) спермий проникает в яйцеклетку, находясь на стадии поздней телофазы. Ядро спермия не проникает в ядро яйцеклетки, а остается лежать рядом с ним. Каждое ядро в дальнейшем начинает подготавливаться к делению обособленно, и объединение их хромосом происходит только на стадии метафазы первого митотического деления зиготы. В оплодотворенной яйцеклетке — в зиготе восстанавливается диплоидное число хромосом. Из зиготы развивается зародыш семени.
После оплодотворения у покрытосеменных растений развивается дополнительный эмбриональный орган — эндосперм, который представляет собой питательное депо зародыша. Начало развития эндосперма вторым оплодотворением. Второй спермий пыльцевой трубки, попадая в зародышевый мешок» сливается с диплоидным ядром центральной клетки зародышевого мешка. При этом образуется набор хромосом: два одинаковых набора хромосом материнского организма и один набор отцовского.
Слияние одного спермия с яйцеклеткой, а другого — с ядром центральной клетки называют двойным оплодотворением. Честь этого открытия, сделанного в 1898 г., принадлежит нашему соотечественнику С. Г. Навашину. Триплоидная природа ядер эндосперма впервые была установлена у скерды (Crepis) М. С. Навашиным в 1915 г.
Образование ткани, питающей зародыш, является особенностью растений. У животных эта функция возложена на запасные питательные вещества яйцеклетки и материнский организм, питающий зародыш через плаценту.
Одна из особенностей оплодотворения у растений, вытекающая из наличия у них двойного оплодотворения, представляет собой явление, называемое ксениями. Этот термин был предложен в 1881 г. В. Фоке. Смысл этого явления заключается в прямом влиянии пыльцы на признаки и свойства эндосперма. Например, имеются сорта кукурузы с желтым эндоспермом (желтые семена) и с белым эндоспермом (белые семена). Если женские цветки белозерного сорта опылить пыльцой желтозерного сорта, то, несмотря на то, что эндосперм развивается на растении белозерного сорта, окраска его будет желтой или бледно-желтой. Следовательно, ядро спермия способно изменить окраску эндосперма, ибо эта ткань так же как и ткань зародыша, гибридного происхождения.
Таков в самых общих чертах процесс оплодотворения у животных и растений. Однако он подвержен приспособительным изменениям в зависимости от особенностей строения половых клеток и биологии размножения, свойственных каждому виду животных и растений.