В том случае, когда от скрещивания особей разных видов удается получить гибрид и он оказывается плодовитым, открывается возможность изучения наследования признаков родителей в потомстве гибридов.
Изменчивость в потомстве гибридов
В том случае, когда от скрещивания особей разных видов удается получить гибрид и он оказывается плодовитым, открывается возможность изучения наследования признаков родителей в потомстве гибридов.
А. С. Серебровский, суммируя накопленный опыт, отмечает, что для гибридов первого поколения наиболее характерно то, что они имеют фенотип, промежуточный между фенотипами скрещиваемых видов; в нем трудно уловить в чистом виде отдельные признаки родителей. Однако в наследовании признаков у гибридов первого поколения существует большое разнообразие: одни гибриды воспроизводят признаки одного родителя, другие — другого; встречается и такое наследование, когда признак у гибрида развит более сильно, чем у любого из родителей; могут появиться и новые признаки, в то время как другие полностью исчезают. Наличие всех этих типов наследования оставляет впечатление промежуточного и мозаичного типов наследования, что и затрудняет изучение генетических закономерностей при отдаленной гибридизации.
Указанные типы наследования теперь во многих случаях понятны. Например, явление расщепления в первом поколении свидетельствует о том, что исходные родительские формы гетерозиготны по гомологичным генам. При скрещивании диких животных, в частности бизона и яка, с крупным рогатым скотом, несущим мутантные доминантные гены белоголовости и комолости, в потомстве наблюдается обычное менделевское расщепление: рецессивные мутации в гибридном потомстве прикрываются соответствующими доминантными аллелями и проявляются у гетерогаметного пола, если они находятся в половой хромосоме.
Анализ наследования при отдаленной гибридизации удается проводить лишь в тех случаях, когда скрещиваемые виды имеют одинаковые наборы гомологичных хромосом, обладающих в гибридной комбинации свойством конъюгировать. В этом случае у гибрида первого поколения в мейозе образуются сбалансированные гаметы и развиваются нормальные половые клетки. Чаще всего это имеет место при скрещивании близких видов, подвидов и разновидностей. У такого гибрида во втором поколении происходит расщепление не только по признакам скрещиваемых форм, но и но признакам, превосходящим даже родовые различия. Так, например, в работе Г. Д. Карпеченко показано, что при скрещивании двух разновидностей культурного шестирядного ячменя (Hordeum vulgare) во втором поколении появляется огромный веер разнообразия признаков, и среди них такие, которые отличаются от родительских. Эти скрещивания раскрывают часть наследственного потенциала изменчивости рода ячменя.
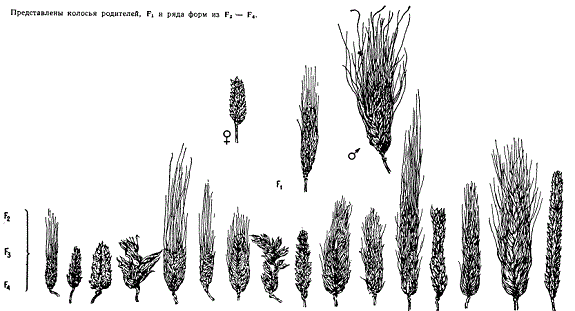
Расщепление гибрида двух географически отдаленных разновидностей ячменя (Hordeum vulgare) — японской (var. Dundar-boyi) и афганской (var. sublatiglumatum)
Такая картина расщепления с большей или меньшей выраженностью обнаруживается в отдельных скрещиваниях, где удается получать плодовитые гибриды.
Роль цитоплазмы в наследовании у гибридов
При отдаленных реципрокных скрещиваниях у межвидовых и межродовых гибридов выявляется роль цитоплазмы в наследовании некоторых признаков.
Это лишний раз свидетельствует о том, что каждому генотипу соответствует своя цитоплазма и что процесс дивергенции видов основывается, не только на различиях в хромосомном аппарате клетки, но и на различиях в цитоплазмы.
Изучение эффекта цитоплазматической наследственности при отдаленной гибридизации проводят несколькими способами, в частности следующими:
- в реципрокных скрещиваниях, где имеется материнский эффект;
- возвратным, или поглотительным (беккроссы), скрещиванием плодовитого гибридного организма с одним из исходных видов;
- путем замещения материнских хромосом отцовскими при контролировании хромосом путем беккросса;
- путем андрогенеза и пересадки ядер.
Уже давно было обнаружено, что от скрещивания организмов разных видов часто развиваются уродливые особи. Аномалии в развитии отдаленных гибридов F1 нередко проявляются в ассиметрии и мозаичности развития. Цитологическое исследование таких гибридов показало, что на разных этапах дробления зиготы происходит элиминация большей части отцовских хромосом, введет, по-видимому, к гаплоидизации клеток. У таких гибридов проявляются в основном материнские черты развития. Поскольку гибриды не достигают половозрелого состояния, дальнейший анализ их оказывается невозможным.
В многолетних исследованиях Ф. Веттштейна по межвидовой гибридизации мхов были получены доказательства того, что наследованная передача некоторых признаков связана с какими-то цитоплазматическими особенностями. При скрещивании видов рода фунария (Funaria), фунарии средиземноморской и фунарии гигрометрической (F. hygrometrica), реципрокные гибриды F1 унаследовали некоторые признаки — тип верхушки листа и жилкование листа — от материнского родителя. Эти гибридные растения в течение восьми поколений подвергались возвратным скрещиваниям с отцовской формой. И хотя ожидалось, что в итоге ее хромосомы материнского вида должны заместиться хромосомами отцовского вида, потомство стойко сохраняло указанные признаки материнского вида.
П. Михаэлис с сотрудниками исследовал роль цитоплазматического наследования у гибридов между различными видами кипрея. Реципрокные гибриды различались по многим признакам, в частности по общей не растения, размерам листьев и цветков, завязей и плодов, фертильности пыльцы. В течение ряда лет гибрид Е. luteum X lirsutum опыляли пыльцой Е. hirsutum, причем в каждом следующем поколении этого возвратного скрещивания в качестве неких форм отбирали растения, наиболее сходные. Уже в четвертом поколении от возвратных скрещиваний получены растения, почти неотличимые по большинству признаков от Е. hirsutum. Такие растения (обозначенные как lh 4) отличали цитоплазму от Е. luteum, а хромосомы, по-видимому, исключительно от Е. hirsutum. Отличия от типичных растений E. lutum, наблюдавшиеся у этой новой формы lh 4, по ряду физических и цитофизиологических показателей (проницаемости и цитоплазмы, устойчивости к грибному заболеванию, отрастания корней и др.), очевидно, должны быть отнесены различий в их цитоплазмах. Далее были проведены контрольные скрещивания растений lh 4 с Е. hirsutum, а затем с Е. luteum. Результаты свидетельствуют о том, что различия между реципрокными гибридами h X 1 и 1 X h почти полностью повторяющимися в реципрокных скрещиваниях с участием новой формы lh 4. У ядра формы lh 4 и Е. hirsutum, по-видимому, почти полностью идентичны, то в данном случае очень четко выявилась роль цитоплазмы в наследовании ряда признаков у межвидовых гибридов, позднее эти данные были подтверждены П. Михаэлисом при повторении беккроссов до 25-го поколения. При скрещивании растений lh 25 с Е. hirsutum в реципрокных направлениях по-прежнему наблюдались различия в зависимости от направления скрещивания.
Один из очень убедительных примеров наследования некоторых признаков у отдаленных гибридов через цитоплазму мы находим в работе К. Корренса с видами бодяка (Cirsium). У Cirsium oleraceum известны как растения с обоеполыми цветками, так и с чисто женскими. При опылении таких женских растений пыльцой с растений, имеющих обоеполые цветки, в потомстве получаются одни женские растения. Скрещивание женских растений С. oleraceum с видом С. canum, который имеет только обоеполые растения, также дает потомство только с женскими соцветиями. Повторение в шести последовательных поколениях скрещивания с С. canum неизменно приводило к появлению только женских растений, несмотря на явное уклонение их по морфологическим признакам в сторону
С. canum. Скрещивание женских растений С. oleraceum с мужскими растениями третьего вида С. palustre, имеющего как женские, так и мужские растения, опять-таки неизменно давало в потомстве только женские растения. Очевидно, женские растения С. oleraceum содержат совершенно специфическую цитоплазму, четко определяющую развитие женских цветков.
Так как в проведенных опытах замена хромосом одного вида хромосомами другого при возвратных скрещиваниях не контролировалась ни генетически, ни цитогенетически, а число хромосом у этих видов достаточно велико, то остается сомнение в отрицании роли последних. При возвратных скрещиваниях и проведении отбора, как это делалось в опытах с кипреем, не исключена возможность сохранения некоторых хромосом материнского вида и детерминирование этих признаков ядерными генами.
Часто наблюдающийся материнский эффект наследования принято объяснять ранней предетерминацией яйцеклеток, генотипом матери, хотя не исключена при этом возможность глубокой видов ой дифференциации плазмы клетки за счет ее самовоспроизводящихся структур.