Кроме рассмотренных внутрихромосомных перестроек, существуют изменения, затрагивающие одновременно две или более гомологичные хромосомы.
К таким перестройкам относятся транслокадии, т. е. обмен участками между негомологичными хромосомами. При транслокациях изменяются группы сцепления генов.
Впервые явление обмена участками между негомологичными хромосомами обнаружено Дж. Беллингом в 1915 г. Он наблюдал 50%-ную стерильность пыльцы 50%-ную дегенерацию зародышевых мешков при определенной системе скрещиваний — бархатных бобов Stizolobium deerigianum. В 1925 г. указанная полустерильность найдена Беллингом также у дурмана (Datura) и объяснена Беллингом как результат сегментных перестроек между негомологичными хромосомами. У дрозофилы транслокация впервые описана Штерном в 1926 г.: она касалась переноса участка Y-xpoмосомы в Х-хромосому. Вскоре транслокации были найдены также у кукурузы по полустерильности початков и пыльцы.
Для того чтобы произошла любая хромосомная перестройка, необходимо осуществление двух процессов: 1) разрыв хромосом и 2) воссоединение оторвавшихся кусков-сегментов с той же самой хромосомой (внутрихромосомные перестройки) или соединение с негомологичной хромосомой (межхромосомные перестройки, или транслокации).
Допустим, что в норме одна пара хромосом несет гены ABCD/ABCD’ а другая пара — гены EFGH/EFGH’. При одновременном разрыве в двух негомологичных хромосомах оторвавшиеся сегменты взаимно обменяются местами: ABGH/ABCd и EFCD/EFGH’.
При этом обменявшиеся участки могут быть равной или неравной длины, и, таким образом, включать равное или неравное количество генов. Такой тип обмена называют взаимной, или реципрокной, транслокацией. Раньше считалось, что существуют и нереципрокные транслокации, однако в последнее время существование их подвергнуто сомнению.
При реципрокной транслокации может происходить объединение двух центромерных участков. В таком случае одна транслоцированная хромосома окажется с двумя центромерами (дицентрическая), а два других сегмента окажутся бесцентромерными и потеряются при делении клетки.
Главной особенностью хромосомных перестроек типа транслокаций является изменение групп сцепления: перемещенные гены войдут в новые группы сцепления и тем самым нарушится сложившаяся система генотипа. Именно на этом основаны генетические методы обнаружения транслокаций. При гибридологическом анализе их устанавливают по изменению групп сцепления и нарушению генного баланса в зиготе.
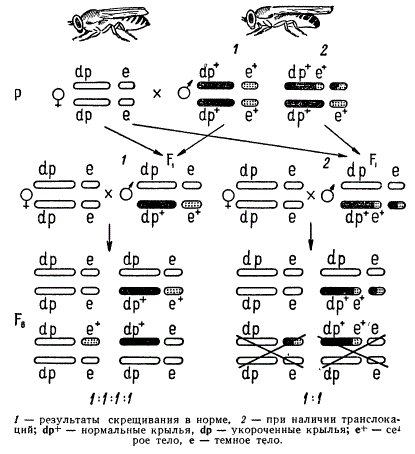
Схема генетического анализа транслокаций
Рассмотрим примеры обнаружения транслокаций у дрозофилы. Для скрещивания берут линию дрозофил, маркированную двумя парами любых рецессивных генов. Например, одна пара генов — укороченные крылья (dumpy) — находится во II группе сцепления, а другая — темно-серое тело (ebony) — в III. С гомозиготной по этим генам самкой скрещивается нормальный самец
![]()
половые клетки которого анализируются на наличие в них взаимной транслокации между II и III парами хромосом.
Из F1 берут самцов, гетерозиготных по указанным генам, и индивидуально скрещивают их с самками из линии с двумя парами рецессивных генов (анализирующее скрещивание):
![]()
В том случае, когда в половых клетках изучаемого нормального самца Р нет реципрокной транслокации между II и III хромосомами, расщепление в потомстве соответствует нормальному отношению 1 : 1 : 1 : 1. В другом случае, когда в одном из сперматозоидов нормального самца произошла транслокация, расщепление изменяется. В результате транслокации II хромосома получила большой фрагмент III хромосомы с локусом е+, а III—малый фрагмент II хромосомы. Сочетание такой мужской гаметы с женской, несущей оба рецессивных гена dp и е, дает гетерозиготу F1 по указанной транслокации dp е / dp+ е+
У самца этого типа образуются четыре сорта гамет. При скрещивании его с самкой, гомозиготной по обоим рецессивным генам, дающей гаметы лишь одного сорта, в потомстве обнаруживается ненормальное расщепление. Так как зиготы, имеющие недостачу или излишек генного материала, не выживают, то особи с фенотипом только dp или только е в потомстве отсутствуют. Нарушение генного баланса приводит к тому, что в Fв появляются не четыре фенотипических класса, а два, т. е. наблюдается расщепление в отношении 1:1.
В последующем генетическими методами, а именно путем изучения результатов кроссинговера, можно локализовать место разрывов в хромосомах. Это же можно производить цитологически.
Реципрокные транслокации появляются, как правило, в гетерозиготном состоянии, так как только одна хромосома из пары гомологичных хромосом обменивается участками с другой, негомологичной хромосомой. Гомозиготы по транслокациям получаются в потомстве от особей, гетерозиготных по ним. При этом гомозиготы иногда оказываются жизнеспособными, если разрывы хромосом и новое положение генов не вызывают летального эффекта.
Большой интерес представляет поведение транслоцированных хромосом в мейозе, так как конъюгация таких хромосом у гетерозиготных особей протекает своеобразно. У гетерозиготы по транслокации в профазе I при конъюгации образуется фигура в виде креста. Она возникает в силу того, что гомологичные локусы, оказавшиеся в разных хромосомах, испытывают взаимное притяжение на стадии зиготены.
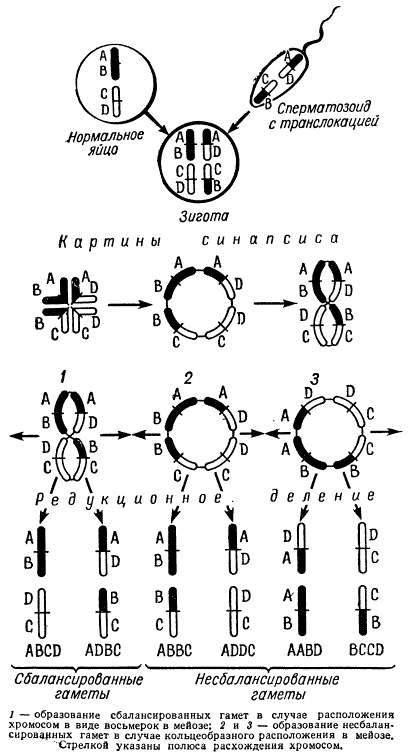
Картины синапсиса и продукты мейоза при гетерозиготных транслокациях
При неразделившихся центромерах в стадии диплотены крестообразные фигуры образуют сложные хиазмы. В диакинезе в силу сползания хиазм от центромер к концам хромосом образуются кольцеобразные фигуры.
Иногда хромосомы кольца, переворачиваясь, образуют фигуры в виде восьмерки. Именно такой тип расположения хромосом и дает жизнеспособные сбалансированные гаметы, так как в этом случае к одному полюсу отходят либо обе измененные хромосомы, либо обе неизмененные. Когда же хромосомы расположены в виде колец в мейозе, образуются гаметы с несбалансированными геномами: в одних — гены повторяются дважды, в других — они отсутствуют, т. е. имеется либо удвоение, либо нехватка их. Следовательно, при равной вероятности всех трех типов расхождения хромосом в мейозе из шести различных сортов гамет, образующихся у организмов, гетерозиготных по одной реципрокной транслокации, только два сорта гамет (т. е. 1/3) оказываются жизнеспособными. Нежизнеспособные гаметы в некоторых случаях можно определить под микроскопом, например стерильные микроспоры у кукурузы, энотеры и др. оказываются сморщенными или пустыми в отличие от нормальных.
Следует отметить, что гетерозиготные транслокации у различных растений, а также у животных дают разную степень стерильности. Эта изменчивость объясняется различием в частоте разных типов ориентации колец на веретене в метафазе I. У кукурузы ориентация хромосом в виде восьмерок происходит примерно в 50% случаев, что приводит к образованию 50% фертильных гамет. У дурмана, пшеницы, ячменя, томатов, энотеры и других растений ориентация в виде восьмерок происходит более часто, что и объясняет большую фертильность их гетерозигот по транслокации.
У ряда высших растений, например у ослинника, пиона, дурмана, колокольчика и др., наличие гетерозиготных транслокаций в генотипе является нормальным состоянием.
Реципрокные транслокации включают две и более пар хромосом. В метафазе образуется кольцо из четырех элементов в результате одной транслокации, кольцо из шести элементов — в результате транслокации между тремя парами хромосом и восьмичленное кольцо — при транслокации между четырьмя парами хромосом.
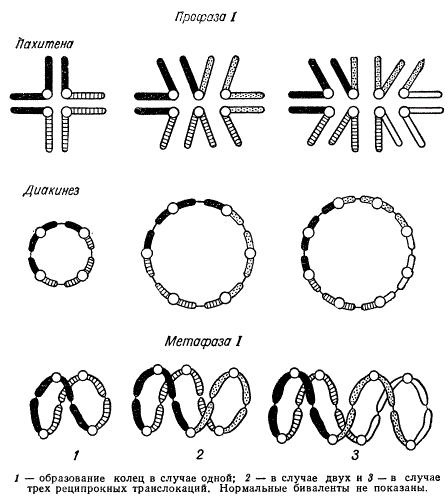
Синапсис хромосом у Oenothera при наличии транслокаций
Из-за образования таких хромосомных комплексов нарушается свободное комбинирование хромосом при редукционном делении, и, как следствие этого, расщепление по признакам будет отличаться от менделевского.
У многих рас ослинника (Oenothera) образование комплексов хромосом является регулярным процессом. Их хромосомы, как это установил Р. Клиленд в 1924 г., в мейозе всегда образуют кольца. Оказалось, что некоторые виды ослинника в мейозе имеют 7 пар нормальных бивалентов, но некоторые расы имеют кольца из четырех и более хромосом, а у вида Ое. muricata и Ое. biennis образуется одно кольцо из всех 14 хромосом.
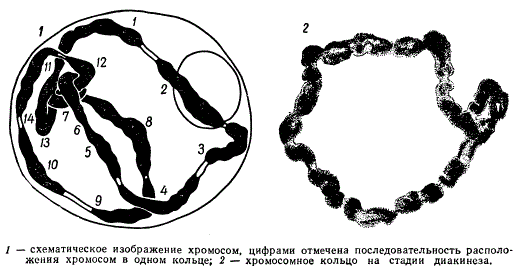
Клеточное ядро Oenothera на стадии диакинеза
Причем в метафазе I один из наборов (7 хромосом) идет к одному полюсу, а другой — к другому полюсу. В этом случае все гены, содержащиеся в каждом из указанных наборов, наследуются вместе, и, следовательно, каждый из потомков всегда получит по два различных комплекса генов.
У Ое. biennis имеются два таких комплекса, которым даны названия rubens и albicans. Каждое из растений этого вида не может содержать два одинаковых комплекса rubens или albicans, а обязательно имеет по одному из них, т. е. является гетерозиготным в отношении сочетания комплексов. Это объясняется следующим образом. В мужских клетках растения образуются два типа пыльцевых зерен — с комплексом rubens и с комплексом albicans, но только комплекс rubens дает нормально развивающуюся и функционирующую пыльцу. Пыльцевые зерна с комплексом albicans не функционируют. В женских половых клетках, наоборот, активным является комплекс albicans и мало активным комплекс rubens. Благодаря различной активности комплексов в женских и мужских половых клетках (гетерогамия) зигота образуется только в одной комбинации rubens + albicans. Следовательно, данный вид энотеры существует на основе комплексной гетерозиготности.
Существует и другой механизм поддержания гетерозиготности видов, основанный на том, что комплексы хромосом функционируют одинаково в мужских и женских половых клетках, но гомозиготное сочетание таких комплексов при оплодотворении ведет к нежизнеспособным зиготам, а гетерозиготное — к жизнеспособным. Такой механизм сохранения кариотипа вида называют иногда сбалансированной летальностью.
Транслокации распространены и среди животных, особенно часто они встречаются у кузнечиков и скорпионов. У бразильских скорпионов Tityus и Isometrus комплексы, включающие транслокации, сходны с таковыми у растений.
Изучение транслокаций представляет не только теоретический интерес, оно может иметь и практическое значение. Мы уже привели пример разделения полов у тутового шелкопряда по окраске грены. Ген, определяющий окраску оболочки грены, в результате транслокации был перенесен из аутосомы в половую хромосому. Таким образом, с помощью транслокации удается изменять группы сцепления у животных и растений в нужном для нас направлении.