Инверсией называют изменение порядка расположения генов в хромосоме, вызванное перевертыванием на 180° большого или маленького участка внутри хромосомы.
Если нормальную последовательность участков с содержащимися в них генами представить в алфавитном порядке АВСД, то при инверсии этот порядок может измениться и стать АСВД.
Для образования инверсии внутри хромосомы необходим разрыв в двух точках, и только при этом условии участок может прикрепиться противоположными концами, перевернувшись на 180°.
Перевертывание концевого участка хромосомы обычно не встречается, вероятно, потому, что конец хромосомы — теломера не обладает свойством воссоединяться с разорванным концом хромосомы. Напротив, разорванным (открытым) концам хромосом свойственна высокая способность к воссоединению. Инверсии часто связаны с рецессивным летальным эффектом, поэтому они не сохраняются в гомозиготном состоянии, и их обычно обнаруживают в гетерозиготе. Однако встречаются и такие инверсии, которые не связаны с летальным эффектом.
Существуют два типа инверсий — парацентрические инверсии и перицентрические инверсии. В случае парацентрической инверсии в хромосоме AB Ο CDEF (где Ο — означает центромеру) расположение генов может быть, например, AB Ο CEDF. В случае перицентрической инверсии последовательность генов ABCD Ο EF изменится на АВЕ Ο DCF.
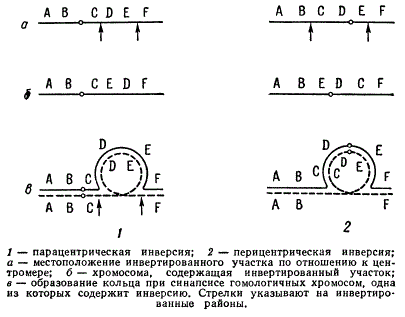
Типы генных инверсий
При гомозиготной инверсии кроссинговер осуществляется нормально. При гетерозиготной инверсии кроссинговер подавляется полностью или частично. Так, например, в гетерозиготной парацентрической инверсии Ο abcdef/Ο AEDCBF хотя и происходит конъюгация хромосом, однако кроссоверных гамет при одинарном кроссинговере не обнаруживается, и это создает впечатление подавления кроссинговера.

Синапсис и перекрест в случае гетерозиотной инверсии в телоцентрической хромосоме и продукты мейоза
В силу нарушения нормального образования продуктов мейоза при скрещивании гетерозиготной по инверсии самки дрозофилы с самцом, несущим только рецессивные гены в гомозиготном состоянии в потомстве не будет возникать кроссоверных особей по данным генам.
Можно видеть, что если происходит одинарный перекрест между хроматидами, то в результате образуются две необычные хромосомы: одна из них без центромеры, а другая — с двумя центромерами. Последняя в анафазе I при полярном расхождении центромер образует хромосомный «мостик», т. е. полярно растянутые хромосомы. Мостик может разорваться в любом месте, в результате чего гаметы получат хромосомы с более или менее значительными нехватками по одним участкам и с дупликациями по другим и поэтому окажутся нежизнеспособными. Нормальные жизнеспособные гаметы могут образоваться только за счет хроматид, не вступивших в перекрест. Так как кроссоверные гаметы оказываются нежизнеспособными, то создается впечатление об отсутствии или подавлении кроссинговера у гетерозигот по инверсиям.
При длинной инверсии, затрагивающей большой район хромосомы (например, в линии СIВ), иногда может обнаруживаться кроссинговер внутри инвертированного участка. В подобном случае кроссоверы возникают за счет двойного перекреста.
Следует заметить, что в гетерозиготных инверсиях по обе стороны от разрывов кроссинговер подавляется в силу отсутствия в этих районах конъюгации хроматид.
В литературе инверсию как подавителя кроссинговера принято обозначать буквой С (начальная буква английского слова crossingover). Это обозначение сохранилось с тех пор, когда подавление кроссинговера приписывалось особому гену в хромосоме.
В отличие от парацентрических инверсий в перицентрических инверсиях ни одиночный, ни двойной кроссинговер не дает ацентрических и дицентрических хроматид, а следовательно, мостов и фрагментов в мейозе. В результате кроссинговера внутри перицентрических инверсий образуются хроматиды с дупликациями и нехватками. Пыльца и яйцеклетки высших растений, имеющие указанные хроматиды, оказываются нежизнеспособными. Церицентрические инверсии могут изменять место центромеры в группе сцепления.
Инверсию можно обнаружить также цитологически в гигантских хромосомах или на пахитенной стадии мейоза.
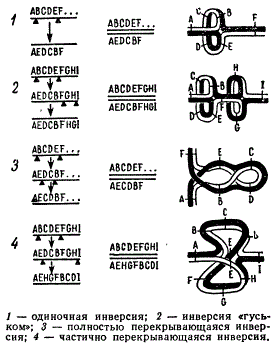
Возможные типы инверсий
Для того чтобы осуществилась конъюгация хромосомы, имеющей инвертированный участок, с нормальной хромосомой у особи, гетерозиготной по инверсии, эти гомологичные хромосомы должны образовать петлю. При этом гены точь-в-точь пригоняются друг к другу. Следовательно, между гомологичными локусами хромосом существует взаимное притяжение огромной силы. Это очень важное явление для понимания физических и химических причин конъюгации гомологичных хромосом в профазе мейоза и ь политенных хромосомах.
Наряду с простыми — одинарными инверсиями в хромосомах встречаются комплексы инверсий, которые могут состоять из нескольких одинарных неперекрывающихся инверсий, называемых иногда инверсиями «гуськом», или сложными инверсиями. В последние включаются полностью или частично перекрывающиеся двойные инверсии.
Итак, в генетическом отношении инверсии характеризуются следующими свойствами:
- в гомозиготных инверсиях, если они не связаны с летальным эффектом, кроссинговер протекает нормально. Причина этого очевидна, поскольку такие гомологичные хромосомы в профазе мейоза могут свободно конъюгировать и обмениваться идентичными участками;
- в гетерозиготных инверсиях, возникающих в двуплечих хромосомах, кроссинговер подавляется только в том плече хромосомы, в котором имеется инверсия; в другом плече подавления перекреста не обнаруживается.
Однако следует иметь в виду, что не всякое подавление кроссинговера может быть отнесено за счет действия инверсии. Существуют такие генные мутации, которые могут препятствовать нормальному синапсису хромосом в профазе мейоза (асинаптические гены), как это установлено у кукурузы, ржи, хлопчатника, дурмана, ячменя, а также у дрозофилы.
Инверсии встречаются в природных популяциях животных и растений, а также могут быть получены в эксперименте, особенно под влиянием ионизирующих излучений и ряда химических веществ. Генетики полагают, что инверсии имеют существенное значение для дивергенции видов. Так, рядом исследований установлено, что отдельные расы внутри вида, например у Drosophila psetrdoobscura, могут различаться по инверсиям. Близкие виды дрозофилы в сходных хромосомах могут иметь обратный порядок расположения генов. Н. Н. Соколов провел цитогенетическое сравнение гигантских хромосом у двух видов дрозофилы (D. virilis и D. littoralis) и у их гибридов, в результате чего выяснилось, что эти два вида различаются между собой шестью инверсиями и несколькими участками, не способными к нормальной соматической конъюгации.
Экспериментально полученные инверсии часто используются в методических целях для создания линий с подавленным кроссинговером.