В природе имеются и электромагнитные системы обнаружения. В результате исследований, проводившихся в СССР и за рубежом, установлено, что некоторые тропические рыбы (мормирус нильский, длиннорыл) обладают органами, имеющими аналогии с устройствами, применяемыми в электроразведке, науке геологического профиля, которая позволяет, изучая электромагнитные поля и электрические свойства горных пород, искать месторождения полезных ископаемых. У этих рыб есть природный генератор электромагнитных разрядов, излучающий электромагнитную энергию в воде. Искажения генерируемого электромагнитного поля, вызываемые появлением в воде различного рода препятствий, улавливаются этими рыбами. Чтобы оценить применимость бионических принципов в электроразведке, во Всесоюзном научно-исследовательском институте разведочной геофизики (ВИРГ) совместно с Институтом экологии и эволюционной морфологии животных АН СССР были проведены специальные исследования.
Установлено, что электрические органы рыб расположены симметрично по бокам тела. Они состоят из набора электрических клеток, имеющих форму пластинок. Лицевая и задняя стороны пластинки обладают электрической полярностью. Если лицевая сторона ориентирована в сторону головы рыбы, то во время разряда голова становится заряженной отрицательно. Электрические клетки-пластинки объединяются в ткани и органы.
Всех рыб по наличию у них электрических органов и электрогенераторных тканей делят на три группы. К первой относятся рыбы, имеющие электрические органы и создающие сильные электрические разряды напряжением больше 10 В, — это электрический угорь, электрический скат, электрический сом и американские звездочеты. Во вторую группу попадают слабоэлектрические рыбы, у которых есть электрические органы, создающие пульсирующие разряды напряжением до 0,5 В (известно около 300 видов таких рыб, например мормириды и гимонотиды). Рыбы, у которых нет специализированных электрических тканей (красноперка, карась, вьюн, окунь и др.), составляют третью группу. Рассмотрим наиболее подробно отдельные виды слабоэлектрических рыб. Они имеют систему, представляющую геофизический интерес.
Электрические органы таких рыб располагаются в хвостовом стебле, замещая там некоторые хвостовые мускулы. Это длинные веретенообразные структуры, лежащие параллельно позвоночнику. Вся электрическая система состоит из элементарных электрических пластин. Например, нильская щука — гимнархус нилотикус (или гимнарх) — имеет восемь электрических органов (по четыре на каждой стороне хвостового стебля). Пластинки располагаются перпендикулярно к позвоночнику по всей длине каждого волокна. Их число в спинном волокне 99, диаметр волокна 4,2 мм, максимальный размер электрической пластинки 1,2 см. Каждая пластинка образуется из 12—20 мышечных клеток. Пластинки всех волокон работают синхронно. В областях хвоста и головы появляются разные заряды, поэтому получаются двуполярные импульсы.
Характерной особенностью электрических органов мормирия является наличие над и под ними удлиненных игольчатых костей. Поэтому при движении рыб орган не изгибается. Пластинки строго перпендикулярны к оси тела. Это обеспечивает постоянную геометрию излучающего диполя и возможность суммировать образующиеся потенциалы.
У многих других видов рыб, как, например, у гимнарха, наблюдается тенденция двигаться с выпрямленным телом или той частью тела, на которой находятся электрические органы. Эти особенности строения хорошо объясняются стремлением к сохранению симметрии электрического поля по отношению к продольной оси тела.
У рыбы африканского слоника, помещенного между электродами, разряд составляет 7—17 В. Электрические органы заканчиваются клиньями из студенистой массы, возникшей из соединительной ткани. Эта масса представляет собой переход от клеток электрических органов к клеткам остальных тканей.
Слабоэлектрические рыбы излучают слабые импульсы электромагнитной энергии. По характеру импульсов-разрядов этих рыб, согласно X. Лиссману, можно разделить на две группы. Разряды первой группы, куда относится и гимнарх, регулярные с относительно большой длительностью импульса (2—10 мс) и частотой изменения амплитуды сигнала в пределах импульса, не зависящей от возбуждения рыбы. Частота следования импульсов — от 60 до 940 в секунду. Разряды второй группы — сложные с меняющейся частотой и амплитудой сигнала в пределах импульса меньшей длительности (до 200 мс). Частота следования импульсов возрастает или уменьшается в зависимости от возбуждения рыбы.
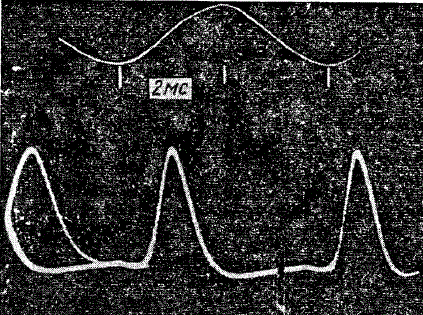
Отдельные импульсы излучения гимнарха, снятые при температуре 28°С
В первой группе наиболее изучен гимнарх, для излучения которого характерны импульсы длительностью 3 мс с интервалами между ними 2,3 мс. Импульсы у гимнарха обнаруживаются и вне воды, если поместить рыбу в воздухе и сделать контактное присоединение электродов к коже. Разность потенциалов между головой и хвостом рыбы достигает сотых долей вольта. При разряде хвост становится заряженным отрицательно относительно передних участков тела. В результате около гимнарха появляется суммарное электрическое поле, эквивалентное полю диполя, силовые линии которого вытянуты вдоль длинной оси тела рыбы. Поле диполя не симметрично — его силовые линии более вытянуты вперед. Это связано с тем, что электрические органы в теле рыбы — элементарные диполи — оканчиваются на кончике хвоста не в одной точке, а в длину простираются на разные уровни. Наблюдения X. Лиссмана показали, что частота колебаний в пределах импульса у одной и той же рыбы не меняется ни при раздражении рыбы, ни при изменении ее физиологического состояния. Изменяется частота разрядов только при изменении температуры. Оптимальная температура для работы электрических органов рыб этой группы 28 °С.
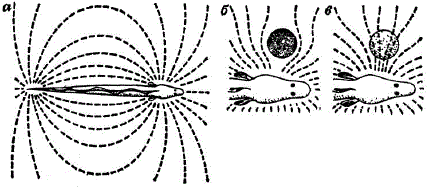
Электрическое поле гимнарха (по X. Лиссману). а — нормальное положение поля вокруг рыбы (вид сверху); б — изменения в электрическом поле, вызванные посторонним предметом (плохим проводником); в — изменения, вызванные хорошим проводником
Рассмотрим разряды второй группы рыб на примере африканского слоника. Разряды слоника состоят из импульсных посылок, заполненных синусоидальным сигналом, частота посылки импульсов изменяется в зависимости от возбуждения рыбы и внешних факторов: температуры, света, солености, вносимых в воду объектов. Длительность каждого импульса составляет от 200—300 до 1 мс, минимальный интервал между ними при высокой частоте заполнения равен 18 мс. Синусоидальные колебания в импульсах содержат частоты от 300 Гц до 20 кГц с максимумами в области 3 кГц. Энергия отдельного импульса составляет 10-6 Вт. При первой фазе разряда рыба создает электрическое поле, которое по форме напоминает поле электрического диполя, также асимметричного, как и в случае, показанном на рисунке, и имеющего большую плотность тока в хвостовой части. Были также произведены специальные измерения, электрических параметров рыбы гнатонемуса как своеобразного эквивалента генератора. Зная основные технические характеристики природного генератора, можно сделать его модель, в которой два электрических органа гнатонемуса заменены диполем с двумя шарами.
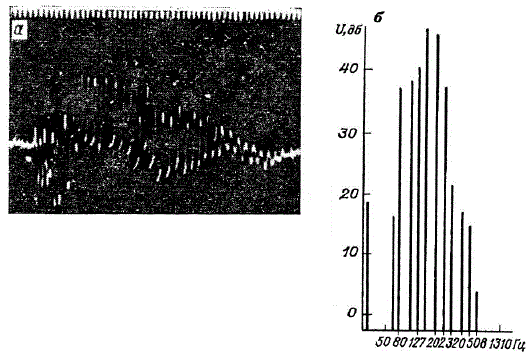
Характеристики электрических разрядов живых объектов (по материалам В. Р. Протасова). а — электрические разряды карася; б — спектры частот электрических разрядов окуня
Электрические разряды у рыб, не имеющих электрических органов, впервые были обнаружены в 1955 г, у миноги, затем у угря В. Р. Протасовым и его коллегами. Разряды неэлектрических рыб сложны по структуре. Амплитуда разрядов не превышает обычно 100—200 мкВ при разносе электродов на длину рыбы. Частоты разряда лежат в области спектра от долей герца до 2 кГц. Разряды неэлектрических рыб представляют собой колебания сложной формы. Длительность разрядов у разных видов рыб варьирует от 5 до 280 с. Пример такого разряда показан на рисунке. Там же дан пример спектра. Для разрядов характерна амплитудная модуляция. Общим для всех исследованных рыб является спектр распределения энергии в разрядах: основная энергия разрядов лежит в области частот от 50 до 800 Гц. В чем-то эти электрические разряды напоминают по форме ультразвуковые колебания аппарата летучей мыши. Особенности излучения живых генераторов электромагнитной энергии интересны и геофизикам.
Напряженность электрического поля у большинства рыб составляет 8—15 мкВ/см на расстоянии 5—10 см от рыбы. Экспериментально измеренное активное сопротивление между двумя электродами, расположенными на близком расстоянии от рыбы, равняется 500 Ом. С помощью электродов зарегистрирован разряд 300 мкВ. Таким образом, мощность разрядов составляет 10-11 Вт, а сила тока 0,6∙10-6 А. Некоторые из разрядов при прослушивании с помощью устройства, содержащего динамик, напоминают «писки — свисты», а другие «морзянку».
Внешние потенциалы могут генерировать и нервные, и мышечные клетки. Однако комплексы этих клеток обладают малой мощностью генерации, так как работают несинхронно. Возникающие разряды состоят из набора импульсов разной длительности, имеют шумовой характер и в результате характеризуются малой мощностью. В случае полной синхронизации в излучении суммарная энергия внешнего поля резко возрастает. Следовательно, можно представить возникновение больших внешних электрических полей при условии синхронной работы нервно-мышечных тканей у неэлектрических рыб.
Исследуя разрядную деятельность неэлектрических рыб и производя параллельный контроль их звучания, В. Р. Протасов обнаружил связь электрических разрядов и звуков. Он доказал, что отдельные электрические разряды часто, но не всегда, сопровождаются звуками. Наиболее часто электрические разряды и звуки у неэлектрических рыб регистрировались при воздействии на рыб ударом палочки, сжатием и т. д., а также в агрессивно-оборонительных ситуациях, так как появление этих разрядов обусловлено деятельностью нервов и мышц. Звуки рыб возникают в результате работы особых акустических аппаратов, основой которых является сокращение мышц. Записи звуков и электрических разрядов ставриды показали, что частотный состав ее разрядов и звуков различается. Максимум звуковой энергии наблюдается на частоте 2,4 кГц, максимум электрической — на частоте 600—800 Гц.
У слабоэлектрических рыб (например, морской звездочет) наблюдается синхронизация в работе акустических и электрических органов. Смысл одновременной генерации разрядов и звуков биологам пока неясен. Синхронизация работы электрических и акустических органов обусловлена деятельностью нервной системы. По-видимому, одновременное функционирование электромагнитных и акустических систем должно повышать надежность локации. Вместе с тем возможно, что здесь мы имеем дело со сложным типом источника: электромагнитным и акустическим. Кстати, и в неживой природе имеются конструкции, которые свидетельствуют о том же.
Действие электрических и магнитных сигналов объясняется следующим образом. Согласно гипотезе X. Лиссмана, органом чувств, воспринимающим электрические сигналы, является боковая линия. У большинства костистых рыб орган боковой линии представлен системой подкожных каналов на голове и двумя идущими вдоль туловища симметричными каналами, в полости которых расположено множество чувствующих органов. Кроме того, тело и голова рыб в той или иной степени покрыты свободнолежащими чувствующими органами — так называемым невромастом. Полости каналов заполнены слизистым содержимым и обычно сообщаются с наружной средой через поры или канальцы. Только у немногих рыб эти поры закрыты мембранами.
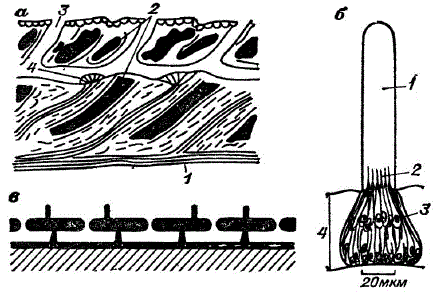
Чувствительные элементы (преобразователи) живых объектов. а — разрез участка боковой линии (1 — нервное волокно, 2 — чешуйки, 3 — канал для притока воды, 4 — рецепторы боковой линии) (по материалам В. Р. Протасова); б — строение чувствительного элемента боковой линии — невромаста костистой рыбы (1 — копула, 2 — сенсорные волоски, 3 — чувствительные рецепторы, 4 — -эпидермис); в — схема строения канала боковой линии (показано в сечении)
Характерной чертой строения невромаста являются бутылкообразные чувствующие клетки, погруженные в самый поверхностный слой кожи. Из каждой клетки наружу выходит чувствительный сенсорный волосок. Несколько сенсорных волосков обыкновенно заключены в массивную капсулу, заполненную студенистым веществом. Капсула является основным органом, воспринимающим механическое раздражение. У видов, ведущих малоподвижный образ жизни, невромасты разбросаны по всему телу в виде обособленных образований. Подвижные рыбы имеют невромасты, расположенные в виде линий на голове или туловище. В результате образуется довольно сложная чувствующая система, слагающаяся из чувствующих капсул, которые расположены на поверхности кожи и в ее глубине. Сенсорные окончания, располагающиеся в глубине кожи, сообщаются с поверхностью через открытые поры.
Особенности строения невромаста послужили основой морфологической структуры для более сложных элементов, именно слуховой капсулы с отолитами у рыб и кортиева органа млекопитающих. Общим в строении отмеченных органов являются желеобразная капсула и чувствующие клетки с сенсорными волосками. К системе органов чувств рыбы, воспринимающих электрические токи, относятся специальные электрорецепторы, расположенные, например, у акул и скатов на голове — ампулы Лоренцини. Гимнарх, используя эти органы, способен улавливать малейшие изменения электрического поля, которые возникают в окружающей его среде (до 0,3 мкА/см2). Кроме того, ряд опытов показал, что гимнарх может различать предметы одной формы, но разной электропроводности.
В настоящее время уже не вызывает сомнения, что магнитное поле влияет на жизнедеятельность живых организмов. Многочисленные эксперименты и наблюдения над животными, человеком и микроорганизмами показали, что колебания напряженности магнитного поля Земли оказывают существенное влияние на развитие практически всех живых существ на Земле. Многолетние наблюдения и результаты сложных биологических экспериментов, подтверждающих этот вывод, были изложены многими авторами во вновь образовавшейся области науки — магнитобиологии. До настоящего времени окончательно не выяснено, каков механизм магнитного влияния.
Одним из наиболее интересных объектов с этой точки зрения являются рыбы. Прежде всего это связано с гипотезами о том, что для многих из них магнитное поле Земли является ориентирующим фактором при миграциях, а также с тем, что они относятся к организмам, у которых обнаружены и уже достаточно хорошо изучены электрорецепторы, или, проще говоря, электроприемники. Следует отметить, что магнитных рецепторов пока не обнаружено ни у одного животного. Более того, многие исследователи (Ю. А. Холодов и др.) пришли к заключению, что рецепция магнитного поля осуществляется либо электрическими рецепторами (например, у рыб), либо непосредственно нервными клетками. Большинство исследователей полагают, что рецепция магнитного поля происходит на клеточном или молекулярном уровне. Многие реакции на магнитное поле оказываются сходными и для микроорганизмов, и для более высокоорганизованных живых существ.
Появляются сообщения и о том, что сейчас находят с помощью магнитометров магнитные материалы, например магнитные кристаллы, в головном мозгу дельфина, тканях головы почтовых голубей, брюшине пчел и т. д. Предполагают, что эти магнитные материалы входят в конструкцию органа чувств, позволяющего живым существам ощущать магнетизм.
Имеется также гипотеза геофизика И. И. Рокитянского о том, что рыбы уходят из области сильного действия магнитотеллурического поля.
Проводившиеся ранее опыты подготовили почву для попытки ответить на еще один вопрос: на какой параметр магнитного поля реагируют живые существа, каковы пороги их чувствительности и как проявляется эта реакция. Это должно было послужить подготовительным этапом к постановке экспериментов по оценке чувствительности к переменному магнитному полю, широко используемому в практике геофизических работ.
Совместными усилиями ВИРГ, лаборатории биофизики АН БССР и Атлантического научно-исследовательского института рыбного хозяйства была разработана методика экспериментов и подготовлена соответствующая измерительная установка.
При экспериментах ставились три основные задачи:
- с достаточной достоверностью доказать наличие или отсутствие реакции рыб на магнитное поле;
- попытаться найти основной «информационный» параметр поля;
- определить реальные чувствительности рыб к магнитному полю.
Эксперименты проводились группой исследователей, в которую входили биофизики, биологи, физики, а также специалисты по радиоэлектронике и геофизике, в том числе и автор. В качестве биологического объекта для изучения влияния магнитного поля на живой организм использовали европейского угря.
Выбор рыб в возрасте около двух лет связан прежде всего с нерестовыми миграциями, при которых угри пересекают Атлантический океан. Угри встречаются на севере во всех странах Балтийского моря и на юге до Балканского полуострова включительно. Такой большой район обитания трудно объяснить только пассивным переносом океаническими течениями, тем более что направление миграции не всегда соответствует направлению определяющих в этих районах течений. Ориентирующими в данном случае могут быть различные факторы, в частности напряженность электрического и магнитного полей.
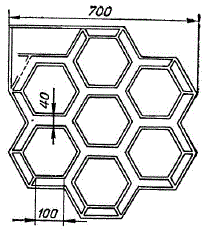
Чертеж лабиринта (по М. И. Глейзеру)
В предположении, что магнитное поле является одним из основных факторов, определяющих направление движения угря во время миграции, было решено выявить, имеется ли зависимость в направлении перемещения рыб от изменений магнитного поля, а если есть, то какие параметры поля являются основными и какова чувствительность к ним. Для изучения характера движения рыб в лабораторных условиях был использован специальный аквариум-лабиринт. Он состоит из шестиугольников и напоминает соты. Чертеж его изображен на рисунке. Ребра шестиугольников являются каналами лабиринта. Всего каналов 36 с номерами от 1 до 36. Выхода из лабиринта нет. Плавание рыб в лабиринте можно рассматривать как процесс блуждания по его каналам. Для изучения влияния внешних факторов на процесс такого блуждания необходимо знать распределение вероятностей переходов из одного канала в другой в зависимости от номера шага, начиная с первого. Шагом при этом назовем движение в одном канале. Таким образом, необходимо установить вероятность перехода из одного канала номер n в другой канал номер n+1. При этом учитывалось, что рыба в канале не разворачивается обратно, так как каналы узки.
Эксперименты проводились в одном из лабораторных помещений, исходя из требований обеспечения максимальной звуко- и светоизоляции и поддержания постоянной температуры около +15 °С. Комната была звукоизолирована прокладками из поролона. Из лаборатории были удалены все металлические предметы, способные привести к искажениям измеряемых магнитных полей. В помещении имелось слабое равномерное освещение бело-голубого цвета с целью имитации освещенности в океане на небольших глубинах, на которых происходит миграция угря.
Аквариум-лабиринт является весьма ответственной частью лабораторной установки. Он не должен быть дополнительным раздражителем, не должен вносить искажений магнитного поля, а его конструкция должна обеспечивать удобную и достоверную регистрацию перемещения рыб. Аквариум-лабиринт размещался внутри рамок, создающих электромагнитное поле. Устройство представляло собой систему излучателей и генераторов, позволяющих скомпенсировать магнитное поле Земли и создать магнитное поле заданной напряженности и направления. Во внутреннем объеме установки оно было однородным или убывающим в пространстве по известному и заданному закону. Излучатели модельной установки представляли собой систему из трех пар колец Гельмгольца, расположенных взаимно перпендикулярно.
Для компенсации магнитного поля на каждую пару колец подавалось постоянное напряжение от трех регулируемых источников постоянного тока. Измерение в зоне нахождения лабиринта производилось магнитометром М-17 с погрешностью ±5γ. Система датчиков располагалась в центре лабиринта и соединялась кабелем с измерительным блоком магнитометра. Магнитометр был установлен на столе оператора, регулирующего магнитное поле, в 2—3 м от модельной установки.
Каждую особь угря помещали в один из отсеков лабиринта, из которого, открывая дверцу, выпускали в лабиринт. Оператор регистрировал переходы рыбы в узлах аквариума, записывая в лабораторный журнал номера соответствующих каналов. Как указывалось, каждый переход в лабиринте имеет определенную теоретическую вероятность. Поэтому можно оценить частоты распределения движения рыб по переходам с заданной достоверностью. По отличию экспериментального распределения от теоретического можно судить о действии внешнего раздражителя, в данном случае магнитного поля.
Для проведения эксперимента использовались 2000 угрей в возрасте около 2 лет, которые были выловлены в Атлантике в момент их миграции к берегам Европы, доставлены в Ленинград и размещены в аквариумах с водой. Во время движения рыбы в лабиринте напряженность поля постепенно увеличивалась от 0 до 2,5 Э.
Такая методика должна была обнаружить порог напряженности, начиная с которого рыбы выбирают переходы, соответствующие их первичной ориентации в магнитном поле, если такая ориентация существует.
Когда было предположительно установлено, что ориентация угрей в магнитном поле существует, для подтверждения этого эффекта, исключения влияния возможных неоднородностей лабиринта и магнитного поля внутри установки, провели серию подобных экспериментов на шести группах рыб по 20—28 в каждой. Направление вектора магнитного поля менялось по отношению к лабиринту на 60°. Для получения возможности статистической оценки получаемых результатов каждый из таких экспериментов повторяли 20 раз, притом на новых рыбах, ранее не запускавшихся в лабиринт.
В результате было установлено, что магнитное поле является не единственным раздражителем, действующим на рыбу.
Видимо, для ряда рыб и других животных магнитное поле играет роль фактора, связанного лишь с рефлексом ориентации живых организмов во время их движения и не имеющего отношения к поиску пищи, уходу от опасности и т. п.
При наблюдении за перемещением угрей в лабиринте на фоне случайного блуждания отмечены особенности движения, выражающиеся в достоверном предпочтении одного, избегании другого и нормальном появлении вдоль третьего направления, которые определялись по странам света. Угол ориентации по отношению к меридиану сохраняется для разных групп угрей и не зависит от конструктивных особенностей экспериментальной установки. Ориентация, с одной стороны, исчезает при компенсации магнитного поля Земли, а с другой — пропадает у угрей, предварительно побывавших в поле электромагнита с индукцией около 0,27 Т, использовавшегося в нашем опыте. По-видимому, ориентация является двигательной реакцией угря на естественное магнитное поле Земли. На основании этого можно сделать предположение о наличии у стекловидных угрей определенной системы, способной воспринимать магнитное поле Земли.
Из экспериментов также следует, что, используя эту систему, угри могут различать страны света в совершенно изолированном помещении. Более того, через функционирование этой системы, угри, по-видимому, способны различать также характеристику, связанную с географической широтой местности. Исследования влияния постоянного магнитного поля показали, что рыбы (угри) реагируют на изменения параметров этого поля. Характер их движения определяется напряженностью и направлением вектора магнитного поля. Основным информационным параметром магнитного поля для рыб является направление такого вектора.
Биологические данные интересны и для геофизического приборостроения. Чувствительность рыб к внешним электромагнитным полям и возможность использования их восприятия для целей локации является одной из наиболее любопытных с точки зрения геофизики особенностей рыб. В связи с тем, что рыбы обитают в хорошо электропроводящей среде (воде), естественно предположить, что они реагируют не только на магнитную, но в первую очередь на электрическую составляющую электромагнитного поля. Как показали эксперименты, такое предположение оказалось обоснованным, так как реакции были примерно одинаковыми в случае задания поля как индукционным способом (с помощью магнитного диполя — рамки с током), так и электрическим способом, например электрическим диполем и двумя электродами. Эксперименты показали также, что характер кривой спектральной чувствительности рыб не меняется под действием шума — электрических помех. По заключению многих исследователей анализатором воспринимаемых электрических сигналов служит мозжечок и продолговатый мозг, представляющие собой высшие центральные отделы органов боковой линии рыб. Высокая степень развития спектральной чувствительности подтверждается, например, способностью нильской щуки выделить полезные сигналы на фоне весьма сильного шума в том же диапазоне частот.
При экспериментах, проведенных В. Р. Протасовым, нильская щука быстро адаптировалась к условиям электрических помех и была способна с неизменной уверенностью электролоцировать находящиеся в воде объекты и реагировать на внешние электромагнитные поля фиксированных частот.
Весьма важный факт при исследовании реакции на магнитное поле был замечен В. Р. Протасовым и при анализе результатов многочисленных зарубежных исследований, проводимых на нильском слонике. В результате изучения этих материалов ученый пришел к выводу, что на воздействие внешнего электрического поля рыба реагирует изменением частоты излучаемого собственного электрического поля.
Приведенное описание экспериментов позволяет сделать предварительный вывод о том, что у электрических рыб восприятие электромагнитных полей в значительной степени связано с рецепцией индуцированных в воде электрических переменных токов, происходящей с помощью известных электрорецепторов.
С целью найти способ обнаружения рыбами подводных объектов и возможности их дифференцирования по физико-химическим свойствам были проведены эксперименты на гимнархе. При этих экспериментах ставились задачи определить следующее: есть ли возможность у гимнарха различать объекты, идентичные геометрически и оптически, но различные по удельной электропроводности; может ли он различать объекты, идентичные геометрически и оптически и одинаковые по электрическому удельному сопротивлению, но с различным химическим составом.
Опыты проводились в бассейне на двух рыбах, у которых вырабатывались условные рефлексы. В качестве определяемых предметов использовались фарфоровые цилиндрические сосуды длиной 15 см и объемом 80 см3. Помещая внутрь сосуда диэлектрики или вещества различной удельной электропроводности, можно было изменять их электрические свойства при неизмененных внешнем виде и электрохимических процессах на границе сосуд — вода. Рыба быстро научилась выбирать сосуды с повышенной удельной электропроводностью (сосуды с водой) — положительный стимул — и игнорировать сосуды с диэлектрическими свойствами (сосуды с парафином, воздухом и с дистиллированной водой). Когда же удельные электропроводности сосудов с дистиллированной, водой и с обычной водой уравнивались, рыба подходила одинаково и к тем, и к другим. Самый Малый объект, электрически отличимый гимнархом от воды, представлял собой стеклянную трубочку диаметром 0,2 см. Таким образом было установлено, что обнаружение объектов происходит электромагнитным способом.
По поводу механизма «локации» высказывалось несколько гипотез. Наиболее вероятная гипотеза была выдвинута X. Лиссманом. Он предположил, что рыба, генерируя электрические колебания, создает вокруг себя поле, эквивалентное создаваемому электрическим диполем.
Термин «локация» ввел для объяснения функций Электромагнитных органов рыб тот же исследователь. Это не радиолокация, когда необходимо иметь антенну, соизмеримую с длиной волны, как условие излучения электромагнитной энергии. Фактически у рыб реализуется чувствительность к искажениям распределения поля в ближней к электромагнитным органам зоне. Распределение потенциалов поля вокруг себя они ощущают с помощью системы электрорецепторов. В результате введения посторонних объектов форма его распределения меняется, а эти изменения напряженности регистрируются. По всей вероятности, последняя гипотеза больше соответствует реальному механизму «локации», хотя и не может объяснить его полностью. При рассмотрении механизма «локации» следует обратить внимание на тот факт, что рыбы (неэлектрические тоже) реагируют на изменения направления электромагнитного поля. Это подтвердили эксперименты, в том числе проводившиеся в ВИРГ. Если рыба сама генерирует электромагнитные колебания, можно предположить, что она способна реагировать и на изменения фазы создаваемого ею внешнего электромагнитного поля, используя в качестве «опорной» фазу генерируемых колебаний. В этом случае при сочетании электрической рецепции по гипотезе X. Лиссмана с фазовой рецепцией биологический объект получает возможность проявить свойства, обнаруженные при описанных выше экспериментах.
Многие рыбы, которые не имеют достаточной возможности видеть объекты, обладают способностью в процессе ознакомления с объектом периодически менять частоту в диапазоне от десятков до 400 Гц. Как правило, при «исследовании» мелких и неэлектропроводных предметов частота выше, чем при исследовании крупных электропроводных объектов. Напряжение, генерируемое при локации, резко отличается от импульсов обороны или нападения. В последнем случае угорь создает серии редких импульсов (по 2—7) большой амплитуды. На основании изложенного можно предположить, что он инстинктивно выбирает «оптимальную» частоту, позволяющую более эффективно «ощущать» исследуемый объект.
Таким образом, у биологических объектов можно предполагать различные механизмы локации, в целом соответствующие нашим геофизическим представлениям. Наиболее изученной их частью является характер генерируемых колебаний, который позволяет высказать предположение, что у рыб имеются механизмы, аналогичные имеющимся в применяемых геофизиками методах, основанных на использовании гармонических и импульсных полей. В отдельных случаях наблюдаются, по-видимому, эффекты, аналогичные проявляющимся в потенциальных полях. У рыб не обнаружено специальных органов, реагирующих на магнитные поля. Считается, что восприятие магнитного поля у животных происходит на клеточном уровне. Можно предположить, что в электропроводной ткани клеток индуцируется электродвижущая сила (ЭДС), которая воспринимается нервными клетками, в том числе и электрорецепторами. Возбуждение живого «датчика» происходит при рыскании рыбы, совершающей резкие периодические отклонения от направления своего движения, при этом действуют электрохимические механизмы электрорецепторов, которые в какой-то степени могут быть уподоблены хемотронным устройствам.
В электроразведке широко применяются различные дипольные установки. В связи с этим геофизический интерес представляет генераторная дипольная установка гимнархуса. Она имеет вид нескольких электрических диполей, конически сходящихся в сторону хвоста и раструбом обращенных в сторону движения. Эти диполи начинаются в районе хвоста и заканчиваются в средней части туловища. По-видимому, система таких диполей создает направленное излучение электромагнитной энергии в сторону движения. Такого вида размещение диполей не используется в известных геофизических электромагнитных установках. Электромагнитные органы биологической установки излучают сигналы сложной формы. Например, длительность импульса может составлять от 300 до 1 мс; синусоидальные колебания содержат частоты от 300 Гц до 20 кГц, минимальный интервал между импульсами 18 мс.
При сравнении частот и продолжительности импульсов биологических органов с используемыми в геофизике обращает на себя внимание их общность. Так, частоты 300 Гц—20 кГц — это основной рабочий диапазон электроразведки для решения поисково-картировочных задач, например в приборах для дипольного электромагнитного профилирования. Импульсы порядка миллисекунд с интервалами в миллисекунды — именно те, что излучает аппаратура по методу переходных процессов. Следует также отметить, что огибающая электромагнитного излучаемого амплитудно-модулированного сигнала является как бы низкочастотным непрерывно действующим излучением работающего органа. По характеру излучения можно предположить, что вторичные поля, излучаемые биологическими электромагнитными органами, в какой-то степени аналогичны вторичным полям, изучаемым в электроразведке. Существенным обстоятельством в характере излучаемых биологических сигналов является динамика. Меняется частота следования импульсов и частота синусоидальных колебаний — все в зависимости от влияния факторов внешней среды. Иными словами, биологические органы периодически меняют характер излучения, добиваясь необходимого оптимума, связанного с внешними условиями.
Приемные диполи расположены с небольшим смещением относительно генераторных. Условно можно считать, что генераторные диполи тяготеют к хвостовой части, а приемные — к передней части туловища. Приемный диполь не один: считая по боковой линии, их два с обеих сторон туловища, они дополнены отдельными скоплениями нервной ткани невромаста. То есть у биологической электромагнитной системы имеется также набор приемных диполей. В пространстве генераторные диполи размещены так, что приблизительно от концов генераторных диполей начинаются приемные. Такого вида установки не известны в геофизике; имеются установки с совмещенными генераторными и приемными диполями или с разнесенными генераторными и приемными рамками. В биологической системе реализуется промежуточный вид установки — с небольшим смещением приемных и генераторных диполей. Дальность действия такой установки составляет, по наблюдениям биологов, несколько метров.
Обращает на себя внимание то обстоятельство, что ультразвуковое излучение летучих мышей также содержит импульсы, выполненные синусоидальными колебаниями. В электромагнитном и звуковом излучении наблюдается одинаковый характер локации, который производится импульсами, выполненными синусоидальными колебаниями. В электромагнитных исследованиях также должно изучаться распределение импульсов, заполненных синусоидальными колебаниями. В геофизических электромагнитных исследованиях изучаются отдельно или импульсные, или синусоидальные колебания. По-видимому, одновременное синусоидально-импульсное излучение позволяет реализовать на практике достоинства как импульсного, так и синусоидального излучения. Действительно, синусоидальное излучение, идущее с перерывом в миллисекунды, воспринимается вольтметром как непрерывная синусоида. Это следует из опытов, проводившихся при излучении двух синусоидальных частот с переключениями; пакет синусоидальных посылок занимал при этом единицы миллисекунд.
При работах по аэроэлектроразведке методом длинного заземленного кабеля кабель длиной 20—30 км запитывали током двух частот, с тем чтобы получить детальную информацию о геологическом разрезе. Осуществлялось это подачей в кабель прерывистыми посылками токов то одной, то другой частоты. Сначала включали ток частотой 976 Гц с продолжительностью посылки несколько миллисекунд, затем 244 Гц, тоже на миллисекунды, и т д. При этом два измерительных устройства регистрировали непрерывно и раздельно сигналы двух гармонических частот — 976 и 244 Гц. Вместе с тем наличие перерыва в посылке позволяло принимать вторичные сигналы при отсутствии первичного поля, например переходные процессы. Такая система излучения электромагнитной энергии наиболее универсальна для решения поисково-картировочных задач и выявления различных по электромагнитным свойствам горных пород и руд.
Для живых организмов подобная система посылок является наиболее гибкой при решении задач навигации, поиска пищи, связи и других в изменяющихся по пути движения электромагнитных свойствах окружающей среды. Удельное сопротивление водной среды, в которой приходится работать биологическим электромагнитным органам, с позиции электроразведки очень низкое. Это единицы, десятки, сотни ом, т. е. условия для электроразведки сложные. По-видимому, для успешной работы в таком диапазоне частот необходимы и выбор оптимальных условий работы, и согласование со средой. Имеются указания на то, что выход биологического генератора согласовывается по сопротивлению с сопротивлением окружающей среды. Обработка полученной информации, управление работой системы, ее подстройка производятся нервной системой. Этот вопрос сегодня изучен недостаточно. Рассмотренная биологическая электромагнитная система, несмотря на небольшие размеры и массу, обладает техническими достоинствами, которые могут найти непосредственное геофизическое применение. В характере излучения и конструкции биологической установки имеется целый ряд привлекающих внимание особенностей: в типе излучения, конструкции диполей, размещении приемных и передающих диполей и т. д.
Заслуживает дальнейшего теоретического и практического изучения строение электромагнитной установки биологической системы: расчет распределения поля, уточнение морфологии биологической системы с точки зрения взаимного расположения генераторных и приемных диполей. Перспективным является изучение спектра и типов колебаний, входящих в состав излучаемого сигнала, а также их изменений.
Следует оценить и электромагнитную систему, состоящую из нескольких расположенных по поверхности конуса диполей, излучающих колебания. Оценка должна содержать элемент биологической работы — тщательное изучение геометрии расположения диполей у разных рыб, элемент физико-математического расчета поля, а в случае необходимости и моделирование. Нужно изучить механизм рецепции о точки зрения определения возможности создания датчиков магнитного и электромагнитного поля на бионическом принципе. Одним из возможных вариантов может быть предположительно ядерно-резонансный датчик, объединенный с хемотронным индикатором сигнала.
Можно считать доказанным, что бионические принципы удастся реализовать в электроразведке. Пока наиболее удобным объектом для изучения биологических электромагнитных систем являются рыбы, поскольку только у них наблюдаются высокоразвитые поисковые электромагнитные органы, позволяющие проводить технические наблюдения. Но этим направлением не исчерпывается возможность изучения датчиков и излучателей биологических объектов. Например, обращают на себя внимание линейные конструкции из электрических клеток-пластин. Их рассмотрение наводит на мысль, что пластинчатые конструкции боковой линии в какой-то степени аналогичны конструкциям пластинчатых антенн. Сравнительный анализ обеих конструкций может иметь технический смысл. Целесообразен более детальный анализ и чувствительных к электрическому воздействию клеток.
С другой стороны, имеются указания на то, что живые системы реагируют и на постоянные поля, т. е. живые датчики достаточно универсальны. Они реагируют на электрические и на магнитные поля по отдельности, а значит, и на совместное действие полей. Различные соотношения электрических и магнитных полей также должны влиять определенным образом на организмы, а механизмы этой более сложной реакции и, следовательно, бионические модели таких механизмов пока не изучены. Одновременное действие электрического и магнитного полей может лечь в основу новых гипотез о чувстве расстояния и ориентации от источника, поскольку отношение электрического и магнитного поля зависит от пеленга. Здесь же скрываются возможности улучшения качества и дальности связи. Но эти механизмы еще ждут своего активного изучения.
Следовательно, анализируя устройство локационных органов чувств живых систем, можно получить полезные технические сведения. Сейчас, когда прошло более двадцати лет официального существования науки бионики, можно сказать, что сделанные прогнозы подтвердились. Бионика стала обычным средством технического моделирования живой природы и позволяет перенести многие из полученных ею результатов изучения живых объектов в мир предметов, создаваемых человеком.
Источник: Г.С. Франтов. Геология и живая природа. (Уровни организации вещества, бионика и геоника, клетки и газово-жидкостные включения). Изд-во «Недра». Ленинград. 1982