Следует поговорить о таком важном свойстве клеток, как возбудимость. В электрическом отношении клетки ведут себя подобно электролитам или смесям электролитов. Вопрос о возбудимости как клеток, так и тканей является предметом изучения электрофизиологии. Клетки и ткани возбуждаются в результате раздражения, природа которого может быть любой: электрической, термической, химической, механической — или носить другой характер.
Энергия раздражения должна превысить некоторый определенный порог. Только после этого наступает раздражение. В опытах раздражителем может быть электрический ток. Величина электрической энергии раздражения Е определяется как произведение напряжения U и тока I длительностью t, или Е = UIt. Наиболее часто раздражение производят импульсом, имеющим прямоугольную форму. Импульс напряжения («толчок напряжения») можно определить, вычислив площадь по кривой, где напряжение представлено в виде графика в зависимости от времени, который называется графиком напряжение — время: Us(t2—t1) = UsΔt.
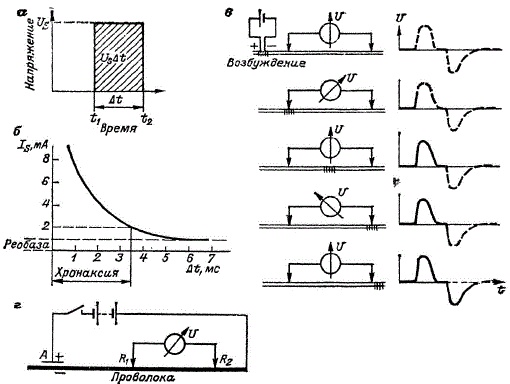
Схемы, поясняющие электрохимическое прохождение импульса. а — прямоугольный импульс напряжения; б — зависимость силы порогового тока от длительности импульса (кривая ток—длительность); в — регистрация «двухфазного потенциала действия», образующегося при распространении волны возбуждения; г — схема проволочной модели нервного возбуждения. Железная проволока находится в концентрированной азотной кислоте. Толчок тока вызывает в модели возбуждение под анодом А. С помощью электродов R1 и R2 регистрируется двухфазное напряжение действия
Толчок напряжения соответственно вызывает в возбудимой системе толчок тока, который является площадью IΔt. Полученная от прямоугольного импульса энергия раздражения будет равна UsIΔt.
Полагая, что падение напряжения Us на биологической системе одинаково, для характеристики раздражения достаточно знать произведение IΔt. Энергию порогового раздражения в электрофизиологии характеризуют значением тока. Для того чтобы пользоваться такой характеристикой, необходимо знать зависимость порогового значения от времени действия тока. В электрофизиологии такую зависимость называют кривой ток — длительность. Она показана на рисунке и очень похожа на геофизическую переходную характеристику.
Существует минимальный ток, называемый реобазой, ниже которого импульс тока, какой бы длинный он ни был, уже не может вызвать возбуждения. Под хронаксией понимают ту наименьшую длительность, которую должен иметь импульс удвоенного по сравнению с реобазой тока, чтобы вызвать возбуждение. На самом деле этот график существенно сложнее. Он строится в логарифмическом масштабе и имеет ряд максимумов и минимумов. И все они связаны со сложными закономерностями живой ткани. Биологи изучают графики сила тока — длительность или кинетический закон порогового раздражения нерва, определяемый зависимостью между пороговой силой и длительностью электрического раздражения. Эту зависимость исследуют на самых различных уровнях организации вещества. В результате была показана универсальная закономерность для живых возбудимых систем, которая наблюдается и при создании импульсного электромагнитного поля над рудными телами. Биофизик У. Ш. Ахмеров обобщил полученные результаты и показал их применимость для технических устройств. Можно сделать и следующий шаг, используя геофизические кривые сила тока — длительность. Мы полагаем, что кривая сила тока — длительность — это общеприродная закономерность, свойственная миру живой и неживой природы. С этой точки зрения кривые сила тока — длительность из мира неживой природы требуют более внимательного изучения, основанного на углубленном представлении о неживом веществе — о минералах и горных породах.
Существенно и то, что для энергии раздражения клетки справедливым является закон суммирования раздражений. Воздействие на тело нейрона определяется суммой воздействий от всех входов и сигналами, действовавшими до этого. Срабатывание нейрона происходит, если воздействие превысит пороговое значение. Тогда на выход нейрона поступает стандартный сигнал. Это означает, что он обладает свойством временного и пространственного суммирования. Последовательным или временным суммированием именуют такое возбуждение нейрона, когда раздражения, меньшие пороговых, следуют через достаточно короткие промежутки времени. Пространственное суммирование состоит в одновременном подведении к двум или нескольким синапсам отдельных раздражений, более слабых, чем пороговые значения. В сумме они могут вызвать возбуждение нейрона.
Интересно, что сразу же после воздействия возбуждающего импульса пороговый уровень нейрона резко возрастает до бесконечности. Значит, никакой вновь приходящий сигнал не заставит его «срабатывать». Такое состояние сохраняется обычно в течение нескольких миллисекунд. Затем пороговый уровень снижается. Если приложить два электрода к наружной поверхности клетки, например к оболочке осевого цилиндра нервного волокна, то под катодом возникнет возбуждение. В теоретическом отношении здесь вводят факторы, способствующие и противодействующие возбуждению. Эти факторы носят на практике электрохимический характер, связанный с закономерностями распределения катионов и анионов. Если подвести к клетке раздражение, превышающее пороговое, то в возбужденном участке клеточной поверхности возникает изменение состояния, захватывающее часть клеточной поверхности и угасающее с увеличением расстояния от места раздражения. На рисунке заштрихованы участки, обозначающие возбужденную зону, отрицательную по знаку относительно соседних участков.
Биофизики постоянно используют в своих работах модели, основанные на аналогиях. Так, отмечено, что между взрывчатой смесью газов, куском железа в азотной кислоте, с одной стороны, и живым нервом, находящимся в состоянии покоя, с другой — существует аналогия, проявляющаяся в том, что обе системы устойчивы по отношению к слабым кратковременным возмущениям. Но когда возмущение превышает определенное пороговое значение, происходит процесс, называемый в случае взрывчатой смеси воспламенением, в случае куска железа — активированием, в случае нерва — возбуждением. Процесс не ограничивается местом приложения возмущения, а распространяется, вообще говоря, на всю систему. На модели нерва ток входит в проволоку, а в натуре ток выходит из нерва. На модели потенциал считается относительно нормального водородного электрода, для нерва берется разность потенциалов между наружной и внутренней частями нерва. Потенциал покоя на модели составляет +1,0 В, для нерва +0,6 В, потенциал действия для модели равен +0,3 В, для нерва —0,035 В, раздражающий толчок тока на модели составляет 2∙10-4, для нерва 2∙10-8 А∙с/см2, плотность тока реобазы на модели 5∙10-3, для нерва 3,5∙10-5 А/см2. Все элементы процесса имеют свои четкие физические характеристики. Модель возбуждения может изучаться при зажигании неоновой лампы с параллельно включенным конденсатором. Система из нескольких таких ячеек с неоновыми лампами моделирует процесс возбуждения. Одна из таких систем, содержащая электротехнические детали, — нейристор X. Крейна — нами была рассмотрена ранее. Схема, по которой производилось изучение возбуждения вдоль железной проволоки, показана на рисунке.
На практике изучение электрического процесса в нервном волокне также, производится с помощью электродов и измерительных приборов. Наиболее часто экспериментаторы изучают очень толстое нервное волокно кальмара. В нервное волокно диаметром 600 мкм вводят четыре серебряные проволочки, через две из них ток пропускают, а через две другие измеряют.
Омическое сопротивление покоящегося нерва кальмара равно 1000 Ом/см2, а его емкость 1 мк Ф/см3. В возбужденной области нерва омическое сопротивление равно 50 Ом/см2, а емкость тоже 1 мкФ/см2. В неживой природе электрические свойства влажных, пропитанных электролитом минералов измеряют в одном из вариантов по той же схеме четырех проволочек-электродов. Толщина двойного слоя электрохимической природы на пропитанном электролитом — влагой минерале составляет 0,01—1 мкм, поверхностное сопротивление — единицы Ом∙м2, емкость — 1—10 мкФ/см2. По-видимому, минералы можно использовать для построения моделей, на которых изучаются электрохимические процессы. Известно, что биологические объекты устроены сложнее геологических. Здесь только обращено внимание на то обстоятельство, что электрические свойства горных пород в значительной степени определяются свойствами электролитов.
С точки зрения электропроводности уместно отметить, что «сухие», вынутые из электролита как белки, так и нуклеиновые кислоты имеют очень высокое удельное сопротивление. Но и обезвоженные, лишенные электролита горные породы также имеют высокое удельное сопротивление (если в них нет добавок электропроводных минералов, например сульфидов или графита).
Биологами Д. Н. Насоновым и В. Я. Александровым была выдвинута теория происхождения биоэлектрических потенциалов. Согласно этой теории, в поврежденной или возбужденной области протоплазмы образуются электролиты, которые при соприкосновении с покоящейся областью протоплазмы обусловливают появление пограничного фазового потенциала. Для появления биоэлектрического потенциала необходим расход энергии, которая образуется за счет обмена веществ. В мякотных нервах миелин образует изоляцию, которая на сравнительно больших расстояниях (через несколько миллиметров) прерывается так называемыми перехватами Ранвье шириной несколько микрометров. Таким образом, практически ток может выходить только в перехватах. Они соответствуют электродам модели. Мы уже видели модель, в которой неустойчивый элемент имитирует неоновая лампочка. Каждая неоновая лампочка соответствует одному перехвату Ранвье. Зажигание одной лампочки вызывает через емкостную связь зажигание последней. Батареи служат для подачи «раздражения».
Изложенное здесь важно для понимания геофизического процесса электрохимических активных горных пород и руд. Не распространяются ли такие волны в природных условиях? Кажется, нет противопоказаний для того, чтобы сродственные волны образовывались и в неживой природе.
Однажды автор принес в лабораторию, исследующую электрохимические нервные процессы, образец — кусочек минерала и попросил изучить электрохимические характеристики минерала. Предварительные результаты показали, что на приборах, которые фиксируют электрохимические процессы, в принципе могут изучаться электрохимические свойства влажных образцов минералов. Так и все изложенное должно заставить задуматься геологов и геофизиков о возможностях открытия нового на стыке биологических и геологических знаний.
Источник: Г.С. Франтов. Геология и живая природа. (Уровни организации вещества, бионика и геоника, клетки и газово-жидкостные включения). Изд-во «Недра». Ленинград. 1982