Полиплоиды, возникающие на основе умножения идентичных наборов хромосом, т. е. наборов хромосом того же вида, называют автополиплоидами.
Автополиплоиды имеют в своем наборе одинаковые геномы. Основное число хромосом (геном) — х соответствует гаплоиду, хх — диплоиду, ххх — триплоиду, хххх — автотетраплойду и т. д.
Автополиплоиды в естественных условиях возникают у растений любым способом размножения. Однако они легче сохраняются у самоопыляющихся растений и при бесполом и вегетативном размножении. Возникшие автополиплоиды в процессе эволюции меняются вследствие мутаций и хромосомных перестроек, которые служат непременным источником разнообразия автополиплоидов. Автополиплоиды широко используются в селекции для получения разных форм со значительной константностью генотипов. Особенно интересны автополиплоиды у форм, размножающихся бесполым и вегетативным путем, так как они могут сохраняться и размножаться в относительно неизменном виде длительное время. При половом размножении автополиплоиды дают также наследственно однообразные формы по числу хромосом, так и по генам, если исходная пара была гомозиготной.
Особенности мейоза у автополиплоидов
Поскольку автополиплоид имеет умноженное число хромосом, комбинирование их в мейозе будет отличным от сочетания хромосом при редукционном делении у диплоидного организма.
В профазе мейоза у диплоидного организма нормально образуются биваленты. У полиплоида, например тетраплоида, в профазе разуются не только биваленты, но и триваленты, и квадриваленты (поскольку могут конъюгировать между собой все гомологогичные хромосомы), и униваленты. При более высокой плоидности возможность конъюгации всех гомологичных хромосом приводит к образованию поливалентов, или мультивалентов.
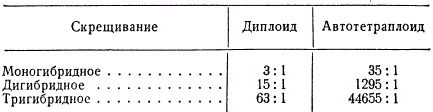
Известно, что если одна из пар хромосом диплоидного организма гетерозиготна по какому-либо гену (Аа), то в результате мейоза обазуются два сорта гамет 2А : 2а. В редукционном делении автотетраплоида ААаа, гетерозиготного по гену А, возникшего а гетерозиготного диплоида, расхождение гомологичных хромосом по полюсам возможно в следующих отношениях 2:2, 3:1, 1:3, 0:4. Если у такого гетерозиготного автотетраплоида ААаа схождение хромосом к полюсам будет проходить регулярно 2 : 2, в этом случае расщепление все же будет отличным от моногибридного расщепления у диплоида. Автотетраплоид, гетерозиготный данной аллели ААаа, образует три типа гамет в отношении АА : 4Аа : 1аа; в F2 расщепление по фенотипу окажется 35 : 1, т. е. будет значительно отличаться от такового у диплоида (3:1). Расщепление 35 : 1 неоднократно подтверждалось в опытах с автотетраплоидными растениями, в частности впервые оно было получено в опытах с дурманом (Datura) при изучении наследования пурпурной и белой окрасок цветка.
При моногибридном расщеплении вероятность появления гомозиготных рецессивных форм у автотетраплоида во много раз меньше, чем у диплоида.
Расщепление по полимерным генам у диплоидных и автотетраплоидных форм
Полиплоидия затрудняет процесс перехода гетерозиготных форм в гомозиготные. Из этого следует, что полиплоидия поддерживает гетерозиготность лучше, чем диплоидный уровень, способствуя тем самым сохранению гетерозиса. Это также указывает на то, что отбор по отдельным генам-признакам рациональнее вести на низком уровне плоидности.
Но, кроме правильного расхождения хромосом, в мейозе у автотетраплоида возможно также расхождение хромосом к полюсам в отношениях и 4 : 0. При этом возникнут гаметы другого сорта, а именно ААа и а, Ааа и А, а также ААаа и 0. Часть таких гамет часто нежизнеспособна. Сочетание таких неполноценных гамет в случае их жизнеспособности может приводить к образованию нежизнеспособных зигот.
Указанное нарушение гаметогенеза является, по-видимому, одной из главных причин того, что автополиплоиды имеют пониженную фертильность. Их плодовитость удается повысить лишь после дополнительной селекции.
Фенотип автополиплоидов
Изучение генетики автополиплоидов представляет особый интерес, так как у них соотношение рецессивных и доминантных аллелей сохраняется таким же, как у исходных диплоидов. Это дозволяет изучать влияние плоидности на проявление признаков.
На первых порах изучения полиплоидии сложилось представление, что полиплоидия у растений обязательно сопровождается увеличением размеров растения и его отдельных органов. Напомню, что одна из первых «мутаций» Oenothera Lamarkiana, обнаруженная Г. де Фризом, была названа «гигантской» (О. gigas) благодаря большим размерам растения. Впоследствии выяснилось, что она является тетраплоидом, имеющим 4n = 48. Однако гигантизм проявляется далеко не у всех полиплоидов, хотя диплоиды по уравнению с гаплоидами всегда несколько крупнее. Полиплоиды, Полученные из гибридных растений разных генетических линий, чаще проявляют гигантизм, чем полиплоиды внутри одной линии. Это указывает на то, что явление гигантизма зависит не только от плоидности, но и от набора, соответствующих генов.
Тетраплоиды у растений, по сравнению с исходными диплоидами, могут иметь большую вегетативную массу, больший размер цветков и вес семян. Гигантизм чаще наблюдается у тетраплоидов перекрестноопыляющихся растений (рожь, гречиха, клевер, турнепс). У, самоопыляющихся растений, например у томатов, полиплоидность не сопровождается гигантизмом. Д. Стеббинс отмечает, что увеличение размеров в результате полиплоидии чаще проявляется щ таких органах, как чашелистики, лепестки, пыльники, семена; у тетраплоидов листья часто толще и шире. Наиболее общим свойством полиплоидов является увеличение размеров клеток. О степени плоидности растения можно судить, например, по размеру зрелых пыльцевых зерен, а также замыкающих клеток устьиц листовой пластинки. Так, размер клеток у полиплоидных форм (Crepis papillaris) находится в прямой зависимости плоидности ядра: при n, 2n, 3n и 4n хромосом размеры клеток равны соответственно 8; 4, 0; 6,0 и 9, 0 (в тыс. мк3). Следующие ряды иллюстрируют увеличение диаметра пыльцевых зерен с увеличением плоидности: в роде Triticum при n хромосом диаметр зерна равен 44,04 мк, при 2n — 51,16 мк, 3n — 55,3 мк, а в роде Rosa при n хромосом — 6,8 мк, 2n — 8,8 мк, 3n — 10,0 мк.
С увеличением размера клеток у полиплоидов иногда наблюдается уменьшение их числа. У мха Funaria гаметофит в норме является гаплоидным, но были найдены и полиплоидные гаметофиты, причем оказалось, что с увеличением плоидности число клеток в поперечнике листа мха Funaria уменьшается, а размер их увеличивается.
С увеличением объёма клеток часто связано изменение ряда их физиологических свойств: относительное увеличение количества воды, уменьшение осмотического давления, изменение содержания различных веществ: белков, хлорофилла, клетчатки, ауксина ряда витаминов и др. Последнее может вызвать вторичные явления устойчивость у полиплоидов к колебаниям внешних заболеваниям и т. д.
У автополиплоидов отмечается нарушение соотношения между процессами роста и развития растения; нередко они оказывают по сравнению с исходными диплоидами менее плодовитыми (по числу семян) и более позднеспелыми. Автополиплоидия проявляется иногда в уменьшении кустистости, например, у ржи.
Плодовитость у автополиплоидов колеблется у разных видов и рас в широких пределах (от 95 до 5%). Одной из причин снижении плодовитости является нарушение нормального расхождения поливалентов в мейозе. Однако существуют специфически действующие мутации, которые влияют на плодовитость полиплоидов, это позволяет при селекции отбирать формы полиплоидов с относительно более высокой плодовитостью. Многие ценные сельскохозяйственные культуры возникли на основе полиплоидии в результате селекции
По исследованиям М. А. Розановой, А. П. Соколовской, О. С. Стрелковой полиплоидные виды в природе часто оказываются наиболее устойчивыми и занимают крайние районы в ареале рода. Более высокие адаптационные свойства полиплоидов, несомненно, определяются их наследственной обогащенностью — более высокой гетерозиготностью.
Устойчивость полиплоидов к воздействию факторов внешней среды объясняется тем, что вероятность проявления у них вредны: рецессивных мутаций значительно меньше, чем у диплоидов. У полиплоидов эти мутации с большей частотой будут оставаться в гетерозиготном состоянии. Так, Л. Стадлер показал что при действии ионизирующих излучений частота появления видимых мутаций больше у диплоидных видов пшениц, чем у тетраплоидных. У гексаплоидных пшениц при тех же условиях мутации вовсе не были обнаружены. Сравнительное изучение коэффициент; изменчивости ряда количественных признаков автотетраплоидной и диплоидной ржи в опытах В. С. Федорова и В. Г. Смирнов; показывает, что коэффициент изменчивости у автотетраплоидной ржи меньше, чем у диплоидной.
Интересно отметить, что «живые ископаемые» растения — представители древних растительных групп оказываются часто полиплоидными. Так, например, среди псилофитовых есть деды, содержащие в наборе 2n = 100 и 2n = 400 хромосом; известно, что у папоротников число хромосом (2n) достигает 500. Большое число наборов хромосом в соматических Щетках имеют также некоторые виды и роды в классах хвощей-плаунов. Полиплоидия является приспособительным механизмом длительного сохранения этих древних форм. По мере развития сравнительной цитогенетики и сопоставления ее данных с палеонтологическими данными о происхождении ныне живущих видов рта новая область исследования (называемая филогенетической цитогенетикой) может многое раскрыть в вопросе эволюции кариотипа при дивергенции видов.
Поскольку скрещивание автотетраплоида с исходным диплоидом ведет к возникновению триплоидных зигот, которые часто оказываются нежизнеспособными или бесплодными, полиплоидия в ряде случаев препятствует скрещиваемости новых форм с исходным полиплоидом, играя роль изолирующего фактора. Так, если посеять рядом диплоидную и автотетраплоидную гречиху, то урожайность семян обоих посевов будет снижена, потому что возникшие от переопыления триплоидные зародыши не развиваются. Подобный эффект наблюдается также при совместном посеве диплоидной и автотетраплоидной ржи. Иногда же, напротив, триплоидные зародыши развиваются в крупные и мощные, но при этом полностью стерильные растения (например, триплоидный арбуз, свекла и др.).
Однако, как показали опыты А. Мюнтцинга, В. В. Сахарова, Р. Жебрака, и других ученых среди популяции автотетраплоидов путем отбора можно получить линии с повышенной плодоносностью. Хорошие результаты селекции автотетраплоидов перекрестноопыляющихся растений дают синтетические популяции, которые составляются из наиболее фертильных линий. Подбор компонентов в виде отдельных линий в синтетическую популяцию и последующее ее размножение позволили В. С. Федорову и Г. Смирнову преодолеть частичную стерильность автотетраплоидов ржи и значительно повысить ее урожайность.
В общем можно сказать, что автополиплоиды по сравнению полиплоидами могут быть: 1) более мощными и плодовитыми, более мощными с пониженной плодовитостью, 3) менее мощными с низкой плодовитостью.
Одной из главных задач в изучении полиплоидии является наследование причин частичной стерильности автополиплоидов перекрестноопыляющихся растений. Благодаря тому, что автополиплоидия сопровождается значительным изменением признаков свойств растения, она оказывается неоценимым резервом исходного материала для селекции.