Генетическое доказательство перекреста хромосом стало возможным благодаря открытию ряда генетических явлений — мутаций, гетерозиготного состояния и сцепления генов.
Первым генетическим объектом, на котором было установлено явление кроссинговера, была дрозофила. Для нее впервые был составлен систематический указатель генов, находящихся в разных хромосомах, и выявлены все группы сцепления.
Рассмотрим один из классических опытов Т. Моргана по перекресту, позволивший ему доказать наследственную дискретность хромосом.
Первые опыты по кроссинговеру
При скрещивании мух, различающихся по двум парам признаков, серых с рудиментарными крыльями (vg — vestigial) b+vg/b+vg и черных (b — black) с нормальными крыльями b vg+/b vg+ в F1 дигетерозиготные особи b+vg/b vg+ по фенотипу оказываются серыми с нормальными крыльями.
На рисунке представлены два направления скрещиваний: в одном дигетерозиготой является самец, в другом — самка. Если гибридные самцы скрещиваются с самками, рецессивными по обоим генам, т. е. производится анализирующее скрещивание b vg/b vg x b+vg/b vg+, то в потомстве получается расщепление в отношении 1 (b+vg/b vg) x 1 (b vg+ x b vg).
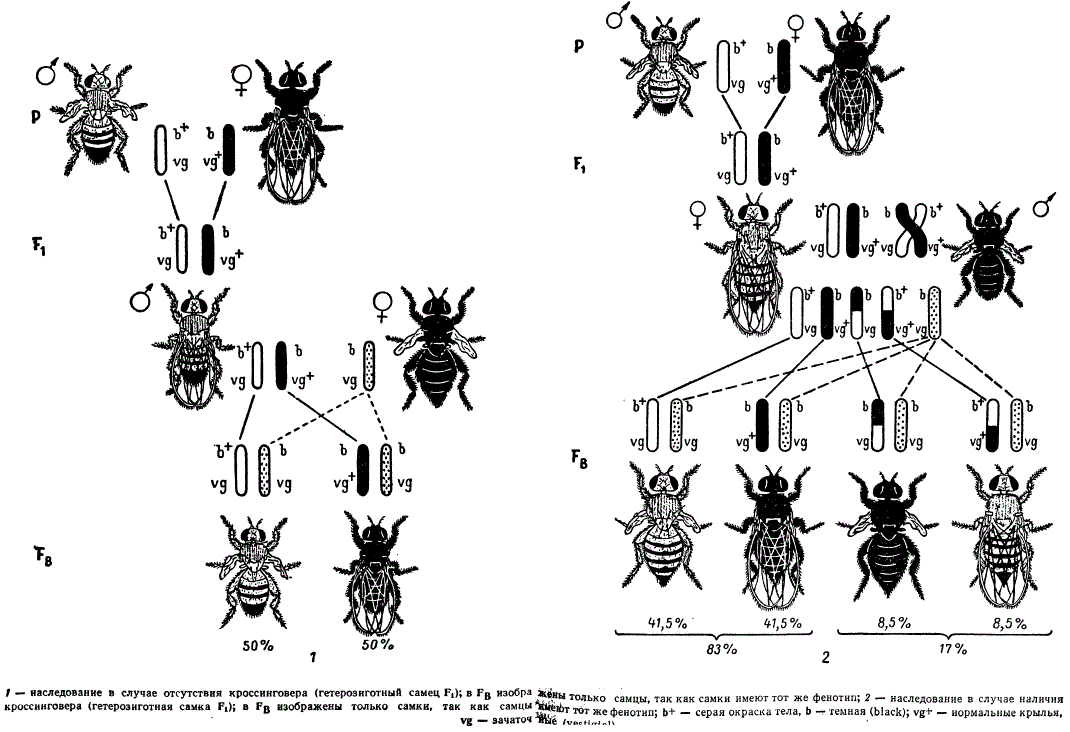
Наследование сцепленных признаков у дрозофилы
Такое расщепление показывает, что данный дигибрид образует только два сорта гамет b+vg и bvg+ вместо четырех, причем сочетание генов в гаметах соответствует тому, которое было у родителей. Исходя из указанного расщепления, следует предположить, что у самца не происходит обмена участками гомологичных хромосом. В дальнейшем выяснилось, что у самцов дрозофилы действительно как в аутосомах, так и в половых хромосомах и норме нет перекреста. Поэтому при описанном анализирующем скрещивании в потомстве восстанавливаются две исходные родительские комбинации признаков: мухи с черной окраской тела и нормальными крыльями и мухи с серой окраской тела и рудиментарными крыльями. При этом они оказываются в равных числовых отношениях независимо от пола. В данном случае мы имеем пример полного сцепления генов, находящихся в одной паре гомологичных аутосом.
В реципрокном скрещивании дигетерозиготных самок с самцом анализатором, гомозиготным по этим же двум рецессивным генам, в потомстве наблюдается иное расщепление. Кроме родительских комбинаций признаков, в потомстве появляются новые типы — мухи с черным телом и короткими (рудиментарными) крыльями, а также мухи с серым телом и нормальными крыльями. Следовательно, в этом скрещивании сцепление генов нарушается. Гены в гомологичных хромосомах поменялись местами благодаря перекресту хромосом.
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными, а не претерпевшие такового — некроссоверными. Соответственно организмы, возникшие в потомстве анализирующего скрещивания от сочетания кроссоверных гамет с гаметами анализатора, называют кроссоверными, или рекомбинантными, а возникшие от сочетания некроссоверных гамет с гаметами анализатора — некроссоверными, или нерекомбинантными.
При анализе расщепления в случае кроссинговера обращает на себя внимание определенное числовое отношение особей разных классов. Обе исходные родительские комбинации признаков, образовавшиеся из некроссоверных гамет, оказываются в потомстве анализирующего скрещивания в равном числовом отношении. В указанном опыте с дрозофилой тех и других особей было примерно по 41,5%. В сумме некроссоверные особи составили 83% от общего числа особей в потомстве. Два кроссоверных класса по числу особей также одинаковы, и сумма их равна 17%.
В качестве другого классического примера сцепления и перекреста можно привести опыт К. Гетчинсона, проведенный на кукурузе в 20-х годах. Были скрещены две гомозиготные линии кукурузы, одна из которых имела зерна с окрашенным алейроном и гладким эндоспермом. Эти признаки определяются доминантными генами c+c+sh+sh+. Другая линия имела рецессивные аллели этих генов c c sh sh, которые определяли соответственно признаки: неокрашенный алейрон и морщинистый эндосперм. Эти пары аллелей находятся в одной паре гомологичных хромосом.
Скрещивание указанных линий между собой c+sh+/c+sh+ x с sh/c sh дает гетерозиготу c+sh+/c sh.
Расщепление при анализирующем скрещивании c+sh+/с sh x c sh/c sh так же, как в опыте с дрозофилой, оказывается не соответствующим независимому поведению каждой пары аллелей. При этом расщеплении количество некроссоверных зерен в початке оказывается 96,4%, а кроссоверных — 3,6%.
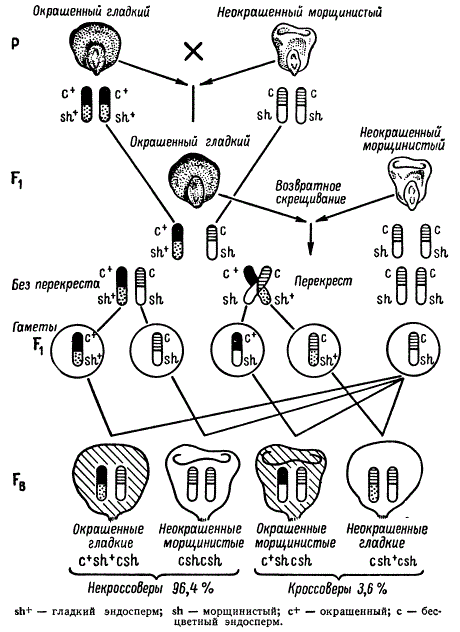
Рекомбинация сцепленных признаков у Zea mays
Результаты опытов на дрозофиле и кукурузе показывают, что действительно существует сцепление генов, и лишь в известном проценте случаев оно нарушается вследствие кроссинговера. Отсюда и следует первое положение о перекресте хромосом, которое гласит, что между гомологичными хромосомами может осуществляться взаимный обмен идентичными участками. Гены, находящиеся в идентичных участках гомологичных хромосом, перемещаются из одной гомологичной хромосомы в другую.
Следовательно, в то время как независимое комбинирование генов, находящихся в негомологичных хромосомах, определяется случайным их расхождением в редукционном делении, рекомбинация сцепленных генов обеспечивается процессом перекреста гомологичных хромосом.
Величина перекреста и линейное расположение генов в хромосоме
Величина кроссинговера измеряется отношением числа кроссоверных особей к общему числу особей в потомстве анализируемого скрещивания и выражается в процентах.
Рекомбинация происходит реципрокно, т. е. между родительскими хромосомами осуществляется взаимный обмен; это и обязывает подсчитывать кроссоверные классы вместе как результат одного события.
Величина перекреста хромосом отражает силу сцепления генов в хромосоме: чем больше величина перекреста, тем меньше сила сцепления. Т. Морган предположил, что частота кроссинговера показывает относительное расстояние между генами: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме, чем реже кроссинговер, тем они ближе друг к другу.
Когда мы указываем, что рекомбинация генов черного цвета тела и коротких крыльев у дрозофилы происходит с частотой 17%, то эта величина определенным образом характеризует расстояние между данными генами в хромосоме. То же самое относится к случаю перекреста хромосом у кукурузы, где 3,6% рекомбинаций показывают частоту осуществленных обменов между двумя участками Гомологичных хромосом.
На основе многочисленных генетических исследований Морган выдвинул гипотезу линейного расположения генов в хромосоме. Только при этом допущении процент рекомбинантов может отражать относительное расстояние между генами в хромосоме.
Одним из классических генетических опытов Моргана, доказывающих линейное расположение генов, был следующий опыт с дрозофилой. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтый цвет тела y (yellow), белый цвет глаз w (white) и вильчатые крылья bi (bifid), были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1160 мух некроссоверных (нормальных и одновременно несущих все три рецессивных признака), 15 мух кроссоверных, возникающих от перекреста между генами у и w, и 43 особи от кроссинговера между генами w и bi. Полученные результаты в процентах перекреста между генами представляли следующее соотношение:
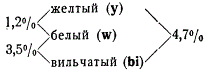
Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами и их последовательного, т. е. линейного, расположения в хромосоме. Расстояние между генами y и bi равно сумме двух одинарных перекрестов между y и w, w и bi.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов вдоль по длине хромосомы строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус.
Одинарный и множественный перекресты хромосом
Приняв положения, что 1) генов в хромосоме может быть много, 2) гены расположены в хромосоме в линейном порядке, 3) каждая аллельная пара занимает определенные и идентичные локусы в гомологичных хромосомах, Т. Морган допустил, что перекрест между гомологичными хромосомами может происходить одновременно в нескольких точках.
Это предположение было им доказано на дрозофиле, а затем полностью подтверждено на ряде животных, растительных объектов и микроорганизмов.
Перекрест, происходящий лишь в одном месте, называют одинарным перекрестом, в двух точках одновременно — двойным, в трех — тройным и т. д., т. е. кроссинговер может быть множественным. Пусть, например, в гомологичной паре хромосом содержатся три пары аллелей в гетерозиготном состоянии: ABC/abc.
Тогда перекрест, произошедший только в участке между генами А и В или между В и С (в разных клетках), будет одинарным. В результате одинарного перекреста возникают в каждом случае только две кроссоверные хромосомы (гаметы), а именно аВС и Abc или АВс и аbС.
Если каждая из этих хромосом будет сочетаться в зиготе с гомологичной хромосомой, несущей все три рецессивные аллели а, b и с, то в потомстве получатся следующие генотипы кроссоверных зигот: аВС/abc и Abc/abc или АВс/abc и аbС/abc.
По проценту кроссоверных классов устанавливают частоту произошедших одинарных обменов между генами А и В или В и С.
В результате одновременного перекреста между А и В и между В и С происходит обмен средним участком хромосомы — двойной обмен. При этом в гетерозиготе возникает новый сорт гамет с кроссоверными хромосомами АbС и аВс, которые выявляются с помощью анализирующего скрещивания. В потомстве появляются зиготы со следующей комбинацией генов: AbC/abc и aBc/abc.
Одинарный и двойной перекресты между гомологичными хромосомами доказываются следующим генетическим анализом. В таблице рассмотрен конкретный опыт на дрозофиле, в котором общее число кроссоверных и некроссоверных особей составляло 521. Этот анализ представлен в общей форме, без указания определенных генов, с тем, чтобы подчеркнуть его принципиальное значение.
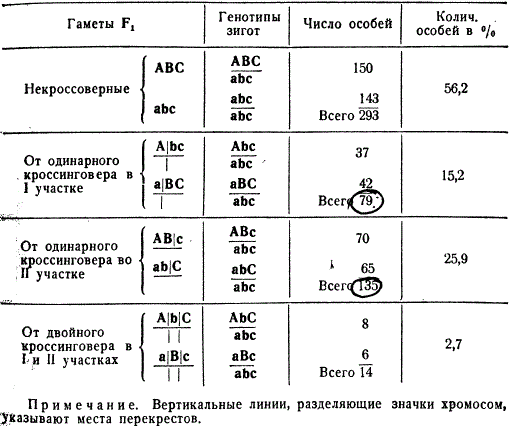
Расщепление в потомстве тригибридов со сцепленными генами при анализирующем скрещивании
Чтобы вычислить процент одинарного перекреста в обоих участках необходимо к одинарным кроссоверам 79 и 135 прибавить число дробей, полученных при двойном перекресте, так как последний произошел как в первом, так и во втором участке.
Рассчитаем процент перекреста между генами A и B: 79 + 14 = 93, 93:521⋅100 = 17,9%.
Один процент перекреста был принят за единицу измерения перекреста, в русской литературе он получил название морганиды.
Следуя тому же методу подсчета перекреста для второго участка — между генами В и С, получим 28,6%, или морганиды. Таким образом, мы определили относительные расстояния между генами: расстояние между А и В — 17,9 и между В и С — 28,6 единиц перекреста, т. е. морганид.
Если правильно, что перекрест есть функция расстояния между генами, то нам легко установить расстояние между генами А и С, так как оно должно быть примерно равно сумме двух частот одинарного перекреста: 17,9 + 28,6 = 46,5. Однако общее число одинарных кроссоверов между генами А и С составляет 214 (79 + 135) особей, или 41,1 морганид, т. е. расстояние между генами А и С, рассчитанное нами ранее, оказалось больше на 46,5 — 41,1 = 5,4 морганиды. Такое расхождение как будто противоречит предыдущему опыту с генами y w bi, где частота перекреста (4,7%) между крайними генами (y и bi) точно совпала с суммой частот перекреста между генами y и w (1,2%) и w и bi (3,5%). Но в том случае гены находятся на близком расстоянии друг от друга, а в примере с генами ABC гены расположены на большом расстоянии друг от друга.
Расхождение в подсчетах объясняется тем, что между далеко отстоящими генами может происходить двойной перекрест, который затрудняет оценку истинного расстояния между генами. Двойной кроссинговер можно не заметить, если расстояние между генами А и С не маркировано третьим геном В.
В случае произошедшего двойного обмена участками внутри хромосом, например, гены А и С останутся на своих местах, и обмен между ними не будет обнаружен. При этом, чем дальше отстоят друг от друга в хромосоме гены А и С, тем больше вероятность двойных перекрестов между ними. Процент рекомбинаций между двумя генами тем точнее отражает расстояние между ними, чем оно меньше, так как в случае малого расстояния уменьшается возможность двойных обменов. Поэтому кроссинговер между генами А и С (41,1%) без учета двойных кроссоверов оказывается меньше, чем сумма единиц кроссинговера между генами А и В, а также между В и С (46,5%).
Для учета двойного кроссинговера необходимо иметь дополнительный метчик, находящийся между двумя изучаемыми генами. В рассмотренном примере таким метчиком является ген В. Определение расстояния от А до С осуществляют следующим образом: к сумме процентов одинарных кроссоверных классов (41,1%) прибавляют удвоенный процент двойных кроссоверов (2,7 X 2 = 5,4%). Удвоение процента двойных кроссоверов необходимо в связи с тем, что каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках. Чтобы вычислить процент одинарного кроссинговера, необходимо величину двойного кроссинговера умножить на 2. В рассматриваемом примере в итоге получается 41,1 + 5,4 = 46,5%, что равняется сумме, полученной от сложения процентов кроссинговера в двух участках: от А до В и от В до С.
Расчет процента кроссинговера между двумя генами можно производить не только на основе данных анализирующего скрещивания, но и по результатам расщепления в F2. Для простоты объяснения предположим, что нам известен процент перекреста между генами А и В и что он равен 20%. Тогда в F1 дигетерозигота AB/ab должна образовать гаметы в следующих соотношениях: 0,4АВ : 0,1Аb : 0,1аВ : 0,4аb (так как кроссоверных гамет 20%, а некроссоверных 80%). В F2 особи, гомозиготные по обоим рецессивным генам, возникают только в результате слияния двух гамет ab с частотой 0,4 X 0,4 = 0,16. В любом случае процент гамет с двумя рецессивными генами у особей F1 определяется как корень квадратный из частоты класса ab в F2, выраженной в долях морганид. В том случае, когда производится скрещивание типа AB/AB x ab/ab, определенная по F2 частота гамет ab, образуемых дигетерозиготой F1 равна половине частоты всех некроссоверных гамет. Если же осуществляется скрещивание типа Ab/Ab x aB/aB, то определенная по F2 частота гамет ab, образуемых гибридом F1 равна половине частоты всех кроссоверных гамет.