А. Н. Северцов предложил очень наглядную схему отношений между ароморфозами и другими типами адаптиоморфозов.
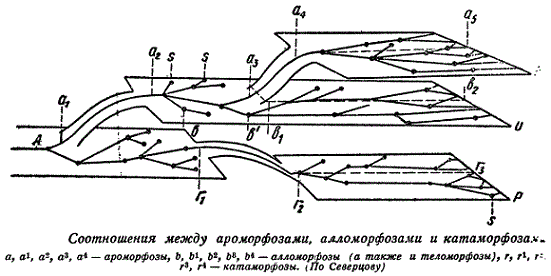
Схема показывает, что в пределах каждой естественной группы за периодом ароморфозов наступает период частных адаптиоморфозов, в ходе которых осуществляется дивергенция (адаптивная радиация) и эволюция. Эти отношения между ароморфозами и другими адаптиоморфозами имеют всеобщее значение и по своей приложимости ко всем группам заслуживают наименования закона А. Н. Северцова. Закон этот может быть выведен из теории естественного отбора, на чем мы должны остановиться, так как вне теории отбора явление адаптиоморфозов остается непонятным. В самом деле — почему в одних случаях наблюдаются ароморфозы, а в других идиоадаптации? Почему за ароморфозами следует период частных приспособлений? Какой фактор управляет этими закономерностями эволюции?
А. Причины и условия возникновения ароморфных преобразований. Легко понять, что ароморфозы, в их морфо-физиологическом выражении, являются результатом наследственных изменений. На почве последних у зародышей возникают новые формообразовательные процессы, приводящие, в их фенотипическом выражении, к возникновению филэмбриогенезов. Достигнув взрослой фазы существования данного животного, филэмбриогенетические изменения развиваются в признаки взрослой формы. Таким образом, ароморфные признаки возникают, как любой другой признак. Однако, качество ароморфных признаков — специфично. Мы видели, что к ним относятся «приспособления широкого значения», не приуроченные к узким частным условиям среды, усиливающие функции органов и позволяющие даже занять новую среду. Таким образом, новые преобразования становятся ароморфозами, если они приобретают роль «приспособлений широкого значения», и если они ведут к интенсификации жизненных процессов, в том числе — к усовершенствованию нервной деятельности, локомоторной функции, функции питания окислительных процессов и т. п. Эта интенсификация физиологические функций сопряжена с соответственными морфологическими преобразованиями. Однако, мы не можем думать, что все эти ароморфные особенности возникли у данного организма одновременно. Различные ароморфные преобразования возникали путем наследственных изменений, влиявших на фенотипические преобразования в отдельных органах. Следовательно, отдельные ароморфные признаки еще не составляют целостной ароморфной организации. Только через комбинирование, и следовательно, через образование новых миксобиотипов могли возникать целые группы ароморфных признаков, сосредоточенных в системе одного организма. Легко понять, что формирование таких организмов с целым «набором» ароморфных преобразований могло идти только через естественный отбор наиболее «ароморфных» миксобиотипов. Представим себе, однако, какое-либо животное, приспособленное к очень частным условиям среды, т. е. с целым «набором» специальных приспособлений. Допустим, далее, что у такого животного возникло какое-либо изменение, имеющее ароморфное значение. Легко понять, что в этом случае новое ароморфное преобразование приведет к некоторому повышению организации. Однако, оно все же останется связанным с признаками специализации в других органах. Поэтому обладатель данного ароморфоза окажется не в состоянии реализовать основную черту ароморфной организации, а именно освоить новую среду обитания и выйти на новый путь эволюции. Для пояснения приведем пример. У орангутана мы видим проявление относительно высоко развитого поведения «разумного типа» (термин А. Н. Северцова). Однако, в условиях высокой специализации к древесному образу жизни, этот несомненный ароморфоз не обеспечивает выходов к дальнейшему повышению организации — конкретно в направлении к человеку. В этом смысле можно утверждать, что «удельный вес» данного ароморфоза не велик. Оранги остаются замкнутыми в питекоидной (обезьяньей) организации и, конечно, филогенетически навсегда останутся в ее пределах. Эта закономерность справедлива для большинства наблюдаемых нами форм. Поэтому число ароморфных этапов, известных нам в истории животного мира, относительно невелико, и они соответствуют узловым этапам эволюционных преобразований, появлению новых типов, классов, определенных отрядов (например, приматы) и т. п. Таким образом, в условиях более или менее специализованной организации «удельный вес» отдельных ароморфозов не велик, так как они входят в систему прочих алломорфных, а тем более тело- и катаморфных преобразований.
Представим себе теперь примитивный организм. Примитивность может иметь разное происхождение. Иногда она — результат вторичных упрощений, например, в условиях теломорфной организации. Такая вторичная примитивность как правило не имеет выходов к целостной ароморфной организации. Основным признаком вторичной примитивности будет узкая специализация (стенадаптивность) соответственных форм и притом форм, как правило, прошедших длинную филогенетическую историю.
Другими чертами отличается первичная примитивность. Последняя, как правило, связана с ясно выраженной мультифункциональностью органов и с филогенетической молодостью группы, следовательно, с отсутствием специализации. Уже самый факт мультифункциональности (пусть, примитивной) органов благоприятен для развития ароморфных преобразований. Если у таких форм возникают ароморфные особенности, то последние сразу приобретают большой «удельный вес», так как не затушёвываются узкой специализацией. Следовательно, экологический эффект (значимость приспособлений в повышении биотического потенциала) ароморфозов полностью проявляется. Поэтому можно утверждать, что примитивная (не специализованная) организация наиболее благоприятна для проявления экологического эффекта ароморфозов. Как правило, каждый новый ароморфный этап в развитии новых групп и начинается с примитивной, менее специализованной организации.
Ветвь рептилий, например, возникла от примитивных амфибий (стегоцефалов), а ветвь млекопитающих от примитивных рептилий (териодонтов). При этом бросается в глаза, что у млекопитающих сохранились некоторые признаки сходства с амфибиями (кожа их богата железами, что свойственно и амфибиям, в черепе имеется два затылочных сочленовных бугра, как у амфибий, число фаланг в пальцах, как у последних, невелико и т. п.). Таким образом, возникнув от рептилий, млекопитающие сохраняют некоторые черты, сходства и с амфибиями. Этот факт свидетельствует о филогенетической близости корней всех трех классов, и следовательно, о том, что примитивность первобытных стегоцефалов была условием возникновения первобытных рептилий, а примитивность последних создавала благоприятные условия для проявления экологического эффекта ароморфозов, послуживших источником возникновения класса млекопитающих. Эти отношения подтверждают значение примитивной организации в развитии ароморфных признаков. Поэтому, при построении филогенетических деревьев, как правило, каждая последующая группа форм естественнее выводится не от «вершины» предшествующей группы, а от ее основания.
Б. Отношения ароморфозов к другим типам адаптиоморфозов. Легко понять, что если бы ароморфозы были единственным путем эволюции, то в каждый данный момент филогенетической истории существовали бы только высшие (для данного момента) формы жизни. Элементарные наблюдения показывают, что этого нет, и что в действительности всегда сосуществуют и низшие и высшие формы.
Сосуществование низших и происшедших от них высших форм было, как мы знаем, объяснено еще Дарвином. Разберем его на примерах отношений между ароморфозами и другими формами адаптиоморфозов.
Было уже указано, что генетическая разнородность любой группы неизбежно возрастает вследствие скрещивания. В ней постоянно возникают новые миксобиотипы. Если группа испытала ароморфные преобразования, и тем самым оказалась поставленной в выгодные условия существования, то численность ее начнет возрастать, а вместе с ней возрастет и интенсивность конкуренции между особями. Выходом из этой конкуренции являются преобразования в новом ароморфном направлении. Однако эти ароморфозы приведут к образованию новой группы форм, появление которых ставит прежние формы в невыгодные условия. Последние будут испытывать давление конкуренции, с одной стороны, с обладателями новых ароморфозов, а с другой стороны, с членами их собственной популяции. Например, конкуренция между стегоцефалами, несомненно, возросла при наличии более высоко организованных котилозавров. Данные палеонтологии показывают также, что древние рептилии оказывали мощное элиминирующее воздействие на амфибий. Ароморфозы стегоцефалов были полезны и давали преимущества в условиях отсутствия других, более высоко организованных форм. Наличие последних (котилозавров) снизило эффект ароморфной организации примитивных стегоцефалов. Примитивная ароморфная организация, в свое время полезная в борьбе за существование, потеряла, в известной степени, свое значение. Мы видим, таким образом, что. с экологической точки зрения, значение ароморфозов, как и всяких вообще адаптаций, относительно. Примитивные стегоцефалы стали на путь вымирания. Однако, они вымерли не целиком. В их популяциях и после ответвления примитивных рептилий развивалась генетическая разнородность и притом, конечно, в разных направлениях. Другими словами, кроме ароморфных миксобиотипов, развившихся в первобытных рептилий, в популяции стегоцефалов развивались миксобиотипы, на основе которых возникали формы, обладавшие частными, и прежде всего, алломорфными признаками. Действительно, возникнув в девоне, стегоцефалы уже в конце его начинают диференцироваться на ряд групп алломорфной организации. Алломорфная организация имела, при наличии рептилий, то экологическое значение, которое утратили ароморфозы примитивных стегоцефалов: она создавала выходы из условий острой взаимной конкуренции между амфибиями и повышала сопротивляемость их элиминирующему воздействию рептилий. Значение алломорфной организации может Сыть показано на следующем примере.
Положим, что в данной области действуют ловкие и быстро передвигающиеся, хорошо вооруженные, чуткие и зоркие хищники. Допустим, далее, что жертва не обладает указанными качествами. В этом случае жертва может быть полностью истреблена. Допустим, однако, что в популяции жертвы появились формы, которые, вместо попыток к бегству, зарываются в землю, т. е. переходят от образа жизни хтонобионта к образу жизни эдафобионта. Такие формы приобретают некоторый шанс на выживание. Их биотический потенциал повышается. Допустим, далее, что среди этих зачинающихся эдафобионтов появились особи с несколько более мощными передними конечностями и немного более длинными и острыми когтями, лучше приспособленными к рытью земли. Они будут зарываться несколько быстрее, чем другие. Хотя эти незначительные изменения ускорят процесс зарывания лишь в небольшой степени, все же это может иметь решающее значение в спасении жизни. Отбор будет идти на выработку приспособлении к быстрому зарыванию. Типичнее эдафобионты среди млекопитающих, например, зарываются очень быстро. В конечном счете отбор пойдет не в сторону повышения организации, а по пути дальнейшей специализации, причем почва становится нишей эдафобионта.
Аналогичное происходило с амфибиями. Так, древние лабиринтодонты (отряд стегоцефалов) сперва развивались как наземноводные формы, а позднее, под влиянием элиминирующего давления рептилий, перешли в воду, т. е. экологически изменились. Хтонобионтностъ оказалась для них неблагоприятной (из-за рептилий), и они возвращаются в стихию их предков, становясь, путем развития соответственных приспособлений, гидробионтами. Как видим, алломорфозы приобрели в этом случае большое значение. Стегоцефалы — предки вымерли, а их потомки (алломорфные группы водных лабиринтодонтов) просуществовали до триаса включительно. Нечто подобное произошло и с другими группами древних амфибий. Став на путь дивергенции, они получила как бы новый импульс к жизни. При этом многие древние амфибии развивались в направлении гиперморфозов. Это давало охарактеризованные выше преимущества. На пути гиперморфозов развился подотряд лабиринтодонтов — Stereospondyli (триас). Однако по мере развития амфибий — такую же (только более пышную) дивергенцию испытали рептилии. Проникновение их в воду положило конец развитию гиперморфных амфибий. В триасе они вымирают. Расцвет рептилий вызвал преобразования амфибий, испытавших дальнейшую дивергентную диференцировку. Часть из них живет в воде (хвостатые амфибии, Urodela), другая ветвь (бесхвостые, Anura) сохранила хвост только в личиночном состоянии, ведя образ жизни, близкий к понятию амфибионтов, третья группа (безногие амфибии, Apoda) стала вести, во взрослом состоянии, жизнь эдафобионтов. Таким образом, стегоцефалы, путем ароморфозов, дали рептилий, а путем идиоадаптаций, выразившихся в развитии частных приспособлений гидро-, амфи- и эдафобионтов — современных амфибий (дивергенция). Эдафобионты, представленные группой Apoda, приобрели облик роющих червеобразных форм, лишенных конечностей и хвоста. Гидробионты (хвостатые) частично сохранили пожизненные наружные жабры, мало подвижные конечности (тритоны) и хорошо приспособленный к плавательной функции хвост. Напротив, бесхвостые, развиваясь в направлении типа амфибионтов, приобрели сильные, подвижные (в особенности задние, прыгательные) ноги. Эта последняя группа пошла по пути завоевания суши, конечно, в пределах возможности, т. е. не слишком далеко от водоемов и во влажных лесах. Таким образом, некоторые бесхвостые, например, жабы (кожа их менее подвергнута высыханию) приблизились к образу жизни хтонобионтов. Некоторые формы приобрели присасывательные диски на пальцах, или клещевидные лапы, охватывающие тонкие ветви и стали дендробионтами, как например, квакша и т. п.
При этом интересы всех этих форм разошлись, конкуренция стала слабее, а биотический потенциал поднялся. На пути идиоадаптаций класс амфибий сохранился в процветающем виде (в особенности бесхвостые) до геологической современности. Конкуренция и отбор явились двигателями этих многообразных идиоадаптивных направлений. Это видно из следующего примера.
В водоемах наша травяная лягушка Rana temporaria испытывает тяжелое давление конкуренций прудовой R. esculenta и озерной R. ridibunda. Последние — хищники и вытесняют травяную лягушку из занятых ими водоемов, поедая молодь. Развитие травяной лягушки пошло по пути размежевания интересов (дивергенция характеров). Прудовая лягушка имеет преимущественно зеленый цвет и держится н: водной растительности, с которой ее окраска гармонирует. Озерная лягушка — типичный гидробионт. Травяная лягушка (по крайней мере молодые особи) обнаруживает переход к жизни лесного хтонобионта, придерживаясь лесной почвы, причем на фоне пестрого лесного настила окраска ее кожи (преобладание бурых пятен) играет роль криптической окраски. Переход к жизни лесного хтонобионта, в особенности использование леса молодыми травяными лягушками, наиболее подверженными нападениям озерной и прудовой лягушек, естественно повысили выживаемость поколения травяной лягушки, улучшили их питание за счет многочисленных лесных насекомых и, следовательно, повысили биотический потенциал вида. Этого не случилось бы, если бы травяная лягушка оставалась амфибионтом. В последнем случае она подвергалась бы значительно более интенсивному истреблению.
На примере эволюции амфибий мы разобрали экологическое значение частных приспособлений и причины сосуществования низших и высших групп организмов. Как видим, на пути развития частных приспособлений (алло- и теломорфозов) данная группа выживает и при наличии более высоко организованных форм. Сохраняя основные ароморфные признаки, она не испытывает, однако, дальнейшего повышения организации. При наличии частных приспособлений, до известной степени гарантирующих от истребления, дальнейшие преобразования не обязательны, и группа, продолжая дивергировать, на долгое геологическое время сохраняет свои основные особенности. Поэтому класс амфибий, возникнув в девоне, продолжает существовать и ныне.
Мы видим, таким образом, одновременно существующими самые различные типы организации — от одноклеточных до млекопитающих.