Выше мы видели, что идиоадаптации и другие формы эволюции приводят к тому, что группа, возникшая на пути ароморфозов, в дальнейшем претерпевает сложное дивергентное развитие.
В ходе последнего осуществляется глубокая экологическая диференцировка или «адаптивная радиация» видов. Виды, входящие в состав естественной группы, захватывают (в пределах общего ареала группы) все те стации, которые она в состоянии захватить по условиям своей организации. Например, амфибии, в пределах доступных им местообитаний, приспособились к жизни гидробионтов, амфибионтов, хтонобионтов (однако, только во влажных местообитаниях), дендробионтов и эдафобионтов. Небольшое число амфибий, например, протей, вошли в состав экологического класса троглобионтов (пещерных форм).
Рептилии, по условиям своей ароморфной организации, смогли занять значительно большую поверхность суши. Если для амфибий все же ведущим экологическим признаком остается амфибионтность, то рептилии являются по преимуществу хтонобионтами. Хорошо защищенная ороговевшая кожа, предохраняющая их от высыхания (мумификации), подвижность и другие ароморфозы позволили рептилиям широко диференцнроваться (радиировать) в пределах хтонобионтного образа жизни. Одновременно через дендробионтность (древесные формы) был открыт путь приспособления к полету и выработке рептилий-авиабионтов, живших в мезозое. Многие рептилии филогенетически вторично завоевали прибрежную полосу суши и воды, а некоторые стали обитателями открытого моря. Они образовали экологический класс вторичных амфибионтов и гидробионтов (вторичными амфи- и гидробионтами их следует называть потому, что сами они — хтонобионты, но произошли от амфибионтных амфибий).
Птицы претерпели подобную же дивергентную экологическую диференцировку. Ведущей для них является авиабионтность, однако и они дали представителей классов хтонобионтов (страусы, новозеландский киви, у нас, в значительной степени, дрофа и др. формы), многочисленных дендробионтов, (дятлы, поползень, пищуха и др. формы), амфибионтов (гусиные в большинстве) и даже гидробионтов (например, пингвины).
Млекопитающие проникли во все стации и образовали многочисленные экологические классы, проявив по условиям своей ароморфной организации необычайно многообразную адаптивную радиацию.
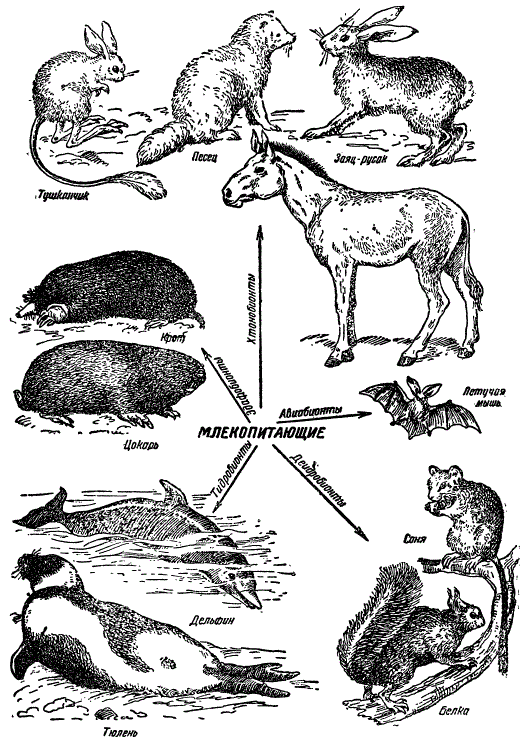
Адаптивная радиация класса млекопитающих.
Показаны некоторые представители жизненных форм — хтонобионты (заяц-русак, тушканчик, песец , эдафобионты (крот, цокарь), гидробионты (тюлень, дельфин), дендробионты (белка, соня), авиабионты (летучая мышь).
Многообразие стаций в природе меньше, чем многообразие естественных групп организмов, почему в одной и той же среде и стации встречаются представители самых различных естественных групп.
Таким образом, по мере образования новых групп позвоночных и их экологической дивергенции (радиации) в самых различных направлениях, каждый экологический класс пополнялся новыми представителями. Заняв одну и ту же стацию и среду, представители различных групп, в процессе филогенетического преобразования, приобрели сходные приспособления (стали конвергентно сходными).
Под влиянием сходного образа жизни различные виды экологически развивались в сходящихся приспособительных направлениях, т. е. конвергировали. Дендробионтность, например, привела к сходному строению конечностей у амфибий, рептилий, птиц и млекопитающих.
Такой путь эволюции и называется конвергентным.
Допустим теперь, что два вида или две группы видов развивались от одного корня сперва в разных направлениях, т. е. в разных условиях, но в дальнейшем, уже после расхождения, проникли в одну и туже среду. Например, обе эти группы были хтонобионтами, а затем стали гидробионтами. Тогда развитие их приспособлений к жизни в воде будет идти параллельно, т. е. они будут проходить параллельные этапы в развитии приспособлений к гидробионтности. Параллельный путь развития одних и тех же приспособлений к одному и тому же образу жизни называется параллелизмом.
Параллелизмы, разумеется, особенно демонстративны, когда они могут быть прослежены во времени. Поэтому они хорошо выявляются на палеонтологическом материале. Примером может служить параллелизм в развитии приспособлений к бегу у вымерших южноамериканских протеротерид (Protherotheridae) — семейство, входящее в состав большой группы копытных, Mesaxonia, куда относятся современные непарнокопытные.
На рисунке показаны конечности трех представителей протеротерид, у которых явственно выражены редукция II и IV пальцев на ряду с интенсификацией III пальца. Этот процесс связан, как и у предков лошади, с нарастающим приспособлением к бегу (этот ряд является, впрочем, ступенчатым, а не филогенетическим). Легко видеть, что развитие приспособленности к бегу шло у протеротерид параллельно такому же процессу у лошадиных предков под влиянием параллелизма в образе жизни.
Возникает вопрос, каковы отношения между дивергенцией, конвергенцией и параллелизмом? С точки зрения дарвиниста, конвергенция и параллелизм являются частными случаями дивергентного развития какой-либо естественной группы.
Прежде всего необходимо подчеркнуть, что понятие дивергенции неразрывно связано с представлением о монофилии. От одного корня на заре развития группы (если только она прогрессивна), как правило, намечаются два основных пути филогенетического развития: путь ароморфозов и путь идиоадаптаций. Однако пути идиоадаптаций вновь расчленяются по разным средам жизни и начинается формирование разных экологических классов (гидро-, амфи-, хтоно-, дендробионтов и т. д.). Возникает специализация (теломорфозы, катаморфозы, гиперморфозы и т. д.). Представители систематически различных классов, на путях этой многогранной дивергенции! («радиации») вливаются в сходные экологические классы, а это открывает пути к возникновению конвергенций и параллелизмов.
Напомним, что во всех этих случаях пути эволюции зависят от естественного отбора. Если, скажем, несколько обособившихся ветвей млекопитающих, в ходе своего эволюционного развития, проникли в воду, и вода стала их нишей, то развитие приспособлений к плаванью, а следовательно, параллелизм и конвергенция превращаются в биологическую необходимость, так как без этих приспособлений становится невозможным успех в конкуренции с другими гидробионтами.
Таким образом, конвергентность и параллелизм в путях эволюции являются естественным следствием конвергентных или параллельных направлений отбора.
Прослеживание филогенетических рядов форм показывает, далее, что во всех группах можно подметить конкретные ряды форм, в которых происходит последовательное развитие определенных признаков. У позвоночных, например, эволюция сердца идет в направлении от однокамерного сердца рыб к трехкамерному сердцу амфибий. У рептилий намечается перегородка между желудочками. У млекопитающих она оказывается полной и сердце становится четырехкамерным. Развитие головного мозга у позвоночных идет в направлении увеличения полушарий переднего мозга и т. д. Подобные явления мы можем проследить по всем системам органов. Мы видели, что у лошадиных предков эволюция шла в направлении развития однокопытности, увеличения роста, моляризации ложнокоренных зубов и т. п. У хоботных наблюдается последовательное увеличение числа гребней на задних коренных зубах, от трех гребней у палеомастодонта до 27 у современных слонов.
Создается впечатление о направленности эволюционного процесса. Это явление было истолковано антидарвинистами, как доказательство «автономического ортогенеза», т. е. что пути эволюции предопределены внутренними автономными факторами развития и носят направленный характер.
Дарвин указывал, что вид, изменяющийся в известном направлении, как правило, будет изменяться в этом же направлении и дальше. Однако, Дарвину была чужда идея ортогенеза в указанном выше смысле. Мысль Дарвина построена на иных основаниях. Если у данного вида возникают некоторые наследственные изменения, то в силу преемственности генотипического развития, в последующих поколениям могут возникать такие же изменения и притом слабее или. напротив, сильнее выраженные. При скрещивании возникают миксобиотипы, у которых эти изменения комбинируются. Допустим, что изменения выражались в редукции II и IV пальцев и увеличении III пальца. Чем определяется этот ортогенез? Ответ ясен: приспособленность к бегу. Рисунок показывает, что в пределах одного и того же рода различные виды (а в пределах вида — различные миксобиотипы обладают различной степенью редукции II и IV пальцев. Например, как видно на рисунке, у Merichippus, primus боковые пальцы длиннее чем у Merichippus pliohippus. Это значит, что последний обладал большей приспособленностью к бегу, чем первый. Отбор и пойдет поэте; линии. Таким образом, ортогенез в данном примере определяется тем что укорочение боковых пальцев связано с лучшей приспособленностью к бегу. Никакой «внутренней» автономной «тенденции», как видим, здесь нет. Развитие ноги лошади в направлении однокопытности определялось не предопределенностью внутреннего порядка, а направлением отбора. В конце концов, о каком ортогенезе внутреннего порядка и «тенденции» в развитии лошади может идти речь, если это последнее представлено пучком форм? Лошадиный ряд от эогиппуса до лошади вовсе не является ортогенезом в смысле предопределенности развития. «Ряд» получается, если мы отвлечемся от всех боковых вымерших ветвей. Это — линия победителе в борьбе за существование, которую мы можем проследите в гуще родословного дерева лошади. Поэтому было совершенно правильно предложено говорить об ортоселекции, т. е. направленности отбора, а не ортогенезе. Направления же отбора зависят от отношений к среде, от того, что данное направление было полезно было естественным результатом конкуренции. Лошадь стала однопалой, потому что в степных просторах однопал ость давала преимущества. Но одновременно было полезнее развитие моляризации зубов так как оно вело к более совершенному перетиранию жесткой злаковой растительности. Полезнее было также иметь больший рост, так как этим достигалось увеличение мощи животного и расширение горизонта. Поэтому отбирались миксобиотипы, совмещавшие все эти признаки.