О справедливости этого положения свидетельствует огромное количество общеизвестных биологических фактов, которые мы лишь напомним, остановившись на главнейших из них.
а) Если органический мир исторически целостен, т. е. имеет монофилетическое происхождение, то следы этой общности его генезиса должны, в той или иной степени, сохраниться в организации любой живой формы. Ближайшие доказательства правильности этого предположения заключены в особенностях организации и функций растительных и животных клеток, т. е. в цитологических фактах. Не задерживаясь на деталях, напомним, что организация животной и растительной клеток принципиально, несомненно, идентична. Особенно демонстративны в этом смысле сходные закономерности митотического деления клеток, протекающего и в животных и в растительных клетках однотипно, по существу — по одной и той же схеме, в порядке одинаковых фаз кариокинеза, совершенно сходного «поведения» хромосом и т. п. Не менее замечательно всеобщее распространение процессов редукционного деления (мейозиса) половых клеток. Процессы мейозиса, по существу, сходны повсюду. Наконец, строение и процессы оплодотворения половых клеток повсюду принципиально однозначны.
б) Сказанное выше может быть, однако, подтверждено и другими данными. Можно говорить не только о принципиальном сходстве в морфологической организации клеток и их делении, но и в целом ряде общих проявлений жизни в них. Явления питания, обмена веществ, роста, возбудимости, размножения и т. п. могут быть сведены к определенным закономерностям, которые иллюстрируются с равным успехом как на животном, так и на растительном материале. Именно на почве этого факта возможно и построение особой дисциплины — общей биологии, всё содержание которой — сплошное доказательство существования признаков и закономерностей жизни, общих для животных и растений. Наличие таких общих для них признаков и закономерностей жизни позволяет защищать взгляд, согласно которому жизнь, как целое, имеет единый «корень» происхождения — только этим и могут быть естественно объяснены упомянутые факты. Мы свободны также от чисто абстрактных, косвенно обосновываемых доказательств существования такого общего «корня». Напротив, мы знаем группу организмов, удовлетворяющую представлениям о нем. Таковы, как известно, жгутиконосцы, совмещающие, как класс, признаки животных и растений. Мы имеем солидные основания полагать, что за счет древних жгутиконосцев возникли два мощных ствола — царство животных и царство растений, и тот факт, что зелёные жгутиконосцы входят как в зоологическую, так и в ботаническую систему, с точки зрения дарвиниста, всегда останется лучшим доказательством единства органического мира — монофилетического происхождения его.
в) От изложенных выше общих данных перейдем к некоторым деталям. Если мы, вслед за Дарвином, можем представить себе органический мир в форме разветвленного «дерева (коралла) жизни» со многими «пучками» боковых ветвей, то каждый такой «пучок» (сравни группы видов, берущих свое начало от общего основания), очевидно, имеет монофилетическое происхождение. Таким образом, мы можем себе представить, что, например, все позвоночные возникли от одного корня. Если это так, то при всем многообразии организации позвоночных, мы должны найти в этой последней следы общности их генезиса. Справедливость этой мысли легко показать.
Одним из доводов в ее пользу может служить изумительное сходство эмбрионов позвоночных животных, на которое было обращено внимание многих исследователей задолго до Дарвина. Сходство между эмбрионами позвоночных носит, при этом, не поверхностный и не случайный характер, а напротив вполне закономерный, касаясь не только общей морфологии тела, но распространяясь также на детали их организации. Нельзя при этом говорить о сходных условиях существования представителей различных классов позвоночных — рыб, амфибий, пресмыкающихся, птиц и млекопитающих и, следовательно, объяснять это сходство аналогичными условиями развития. Напротив, сходство между эмбрионами сохраняется вопреки глубоким различиям в условиях развития. Ту же картину находим мы и за пределами класса позвоночных. Едва ли, например, возможно говорить об одинаковых условиях развития морских, пресноводных и особенно паразитических ракообразных. Однако многим из них, независимо от среды обитания, свойственна личиночная стадия — науплиус. Подобные явления сходства эмбриональных стадий развития, в пределах естественных групп, могут получить только одно рациональное объяснение, не требующее каких-либо натяжек, а именно: это сходство есть прямое выражение родства.
г) Перейдем теперь к сравнению взрослых фаз развития. Рисунок показывает, что с наступлением более поздних стадий развития эмбрионов сходство между ними затушёвывается накоплением нарастающих различий.
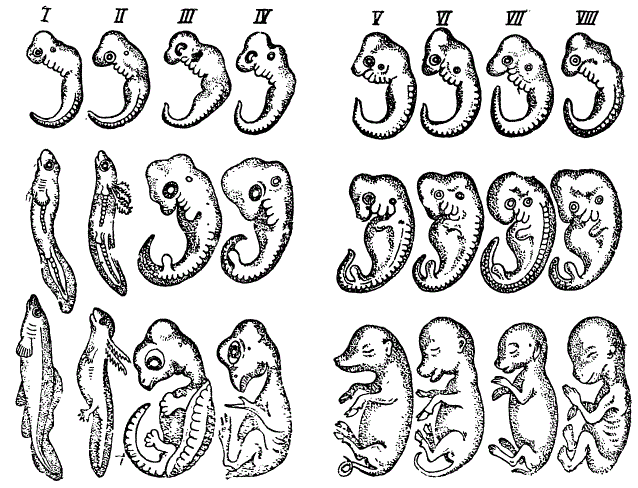
Явление сходства между эмбрионами позвоночных. В вертикальных рядах: I — рыба, II — амфибия, III — рептилия, IV — птица, V—VIII — млекопитающее. Сходство очень велико на ранних стадиях развития (верхний ряд) и менее выражено на более поздних стадиях (онтогенетическая дивергенция, или обособление по Бэру). Из Роменса
Сходство между взрослыми формами еще менее полно. Однако, внимательное сравнительна: анатомическое исследование обнаруживает, что и взрослые формы одной и той же естественной группы сохраняют черты глубокого принципиального сходства. Различия между ними покоятся на сходной основе.
Это факт явственно выступает, если мы учтем, что, как правило, многообразным формам одной естественной группы организмов свойственен общий или единый план строения (план композиции по Кювье).
Под планом строения понимается определенный тип организации. Например, позвоночным животным свойственен общий тип, или план строения. На рисунке показано единство плана строения скелета лошади и человека.
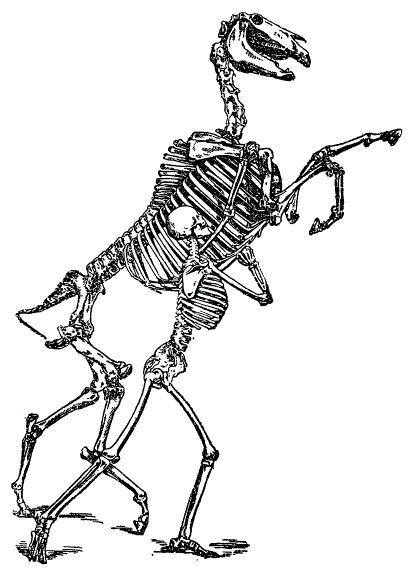
Единство плана строения лошади и человека (Из Мэтью)
С бросающимся в глаза постоянством сохраняются составные части скелета: осевой скелет (позвоночный столб), череп, передний (плечевой) и задний (тазовый) пояса конечностей, соответствующие им передние и задние конечности. Следовательно, в основе представленных скелетов лежит единство общего плана их организации. Явление это распространяется на все системы органов, как наружных, так и внутренних.
Аналогичные отношения могут быть установлены и в пределах других типов животного, а равно и растительного царства. Явление единства плана строения требует причинного объяснения. Учение Дарвина дает простой ответ: наличие единства общего плана строения у многообразных видов одной естественной группы объясняется единством их происхождения.
Ценность этого ответа, однако, ограничивается тем, что единство плана устанавливается только для отношений внутри естественных групп, в конечном счете — для видов одного типа.
Разумеется, не может быть и речи об едином плане, например, для всех типов животного царства. Однако исторический взгляд на органическую природу приводит к необходимости установления связей между отдельными типами и, следовательно, между отдельными планами строения. Возможно ли связать отдельные типы? Да, возможно. Отдельные типы не являются замкнутыми системами организации. В зоологии и ботанике можно найти большое число примеров, из которых видно, что высшие типы организации выводятся из примыкающих к ним низших типов. Иногда связи между отдельными типами необычайно наглядны. Таковы, например, отношения между кольчецами (аннелиды, Annelides) и членистоногими. Проницательный Кювье не без основания соединял их в одну ветвь — членистых (Articulata). Так, представителям обоих типов свойственно сегментное строение тела, каждому сегменту (кольцу) тела соответствует всегда лишь одна пара придатков (пароподий у аннелид, членистые ножки у членистоногих), центральная нервная система в пределах обоих типов представлена брюшной нервной цепочкой, и там и здесь имеется окологлоточное нервное кольцо. Эти черты, указывающие на близость между обоими типами животных, получают еще более глубокий смысл, если мы вспомним замечательную организацию первичнотрахейных (Protracheata). У представителей этой группы ещё сохраняются типичные сегментарные выделительные органы (как у кольчецов), конечности нерасчленены (как параподии кольчецов), стенка тела (как у кольчецов) представлена типичным кожно-мускульным мешком, а с другой стороны, у них имеются примитивные трахеи и ротовые конечности — признаки, свойственные членистоногим.
Аналогичным образом, т. е. сравнительно-морфологическим методом, устанавливается наличие связей и между другими типами животного царства. Исследователь, занятый этими вопросами, стремится составить общее генеалогическое древо, в котором устанавливаются связи между всеми типами животного царства.
Аналогичное генеалогическое древо строится и для растительных групп. Правда, связи между типами часто неясны. Однако с точки зрения дарвиниста это вполне понятно. Неясность сходства между представителями разных типов — результат утраты сходства, т. е. результат изменяемости.
Единство плана строения в пределах естественной группы организмов, как выражение их филогенетического родства, носит при этом детальный характер, проникая в структуру и общую архитектонику отдельных органов и частей тела. Это обстоятельство еще резче и убедительнее подчеркивает, что сходство есть следствие и выражение родства. Сказанное видно из того факта, что, например, животным, обладающим сходным планом строения, свойственны гомологичные органы. В широком смысле слова, под гомологией понимается сходство между органами одинакового происхождения. Гомология наблюдается в различных формах. По Гегенбауру (1874) следует различать общую и частную гомологию. Мы здесь рассмотрим сперва последнюю, так как она охватывает отношения между органами одинакового происхождения, принадлежащими разным организмам, построенным по одному плану.
В этих случаях факты явственного родства между различными формами одной естественной группы выступают очень ярко. На рисунке показаны примеры полной специальной гомологии — правые передние конечности ряда позвоночных.
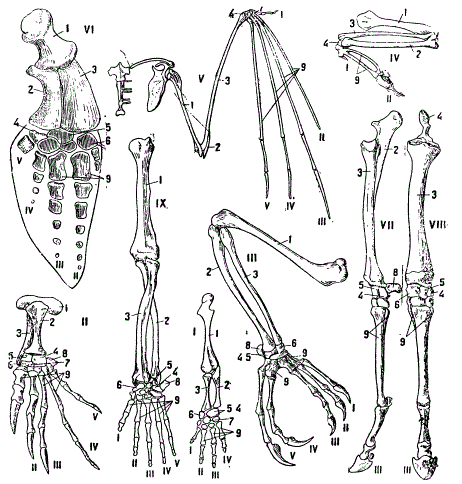
Полная специальная гомология. Крупные римские цифры обозначают: I — нога саламандры, II — нога трехкоготной черепахи, III — нога хамелеона, IV — нога (крыло голубя, V — крыло летучей мыши ( Vespertilio murinus), VI — нога (плавник) дельфина (Phocaena communis), VII—VIII — нога лошади, IX — нога обезьяны. Повсюду: I — плечо, 2 — локтевая кость, 3 — лучевая к., 4 —ulnare, 5 — intermedius, 6 — radiate 7 — centrale, 8 — pisiformis, 9 — пясть. Римские цифры у пальцев показывают пальцы от первого до пятого (По разным авторам, преимущественно из Бючли)
Всюду, как видим, сохраняется один и тот же план строения и притом не только для более крупных частей передней конечности (плечо, обе кости предплечья), но и для деталей организации пясти и запястья. Эти явления ярко свидетельствуют о родстве и не могут быть объяснены ничем иным, например, сходным образом жизни, так как подобраны конечности форм, ведущих различный образ жизни.
Сказанное выше по поводу передней правой конечности с той же убедительностью проявляется при сравнении решительно всех частей тела, общей топографии органов и т. п. Это обстоятельство в высокой степени облегчает приведение многообразия форм в известную систему, в пределах которой, например, сходные виды образуют одну естественную группу. На рисунке показаны два вида нематод из семейства Chromadoridae.
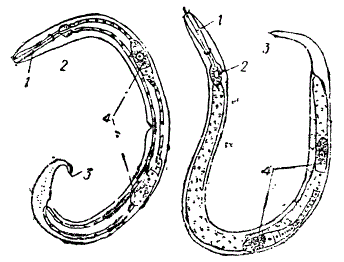
Две нематоды из сем. Chromadoridae. Слева морская форма, справа — пресноводная. Несмотря на различие в среде обитания, сохраняется тот же план строения. 1 — пищевод, 2 — бульбус, 3 — концевая трубка выделительных хвостовых желез, 4 — яичник (По Шурманс-Стекговену и де Ману)
Прозрачность кутикулы позволяет видеть сходство их анатомической организации — один и тот же общий план ее, который сохраняется при всех частных различиях между ними. Явление родства выступает совершенно ясно, особенно, если учесть, что представленные виды нематод весьма различны по местообитаниям (пресноводная и морская). Глубоко различная среда их местообитания и различия в образе жизни обусловили их видовые различия. Однако внешняя среда, на данном отрезке времени, не отразилась на общих им основных признаках. Эти последние накладывают на них общую печать. Каждая естественная группа организмов характеризуется общей для всех входящих в ее состав видов системой признаков, развивающихся у всех особей этих видов, независимо от условий существования.
Таким образом, например, в любой части света, в любых климатических условиях, на севере, в тропиках, в условиях пустыни и на болоте любой вид злаков, характеризуясь своими видовыми признаками, составляющими его особенные черты, в то же время будет обладать признаками, общими для всех злаков.
Каковы же причины этого замечательного явления, определяющего границы естественной группы? Почему известная естественная группа видов (например, злаки), при всем их частном многообразии, сохраняет в любом климате общие им всем черты, по которым тотчас устанавливается, что, например, данное растение — злак, или что данная нематода принадлежит к сем. Monhysteridae и т. п.? Совершенно очевидно, что такое стойкое сохранение известной системы общих сходных признаков, не исчезающих при любых условиях существования, — прямое следствие общности их происхождения. Этот простой и ясный ответ подчеркивает, что любая естественная группа берет начало от общего корня. Отсюда и понятно, что виды одной естественной группы должны иметь известное число общих им всем признаков.
д) До сих пор мы имели дело с фактами морфологического порядка. Однако, каждый организм имеет конкретную морфо-физиологическую характеристику, и форма любого органа и организма в целом всегда неразрывно связана с их функциями, с их физиологией. Очень большое число физиологических и биохимических фактов свидетельствует о единстве, монофилии отдельных естественных групп. Напомним и здесь некоторые примеры.
Известно, например, что гемоглобин и близкие ему пигменты, входят в состав крови очень многих животных. Кроме позвоночных, близкие к гемоглобину пигменты имеются у немертин, пиявок, полихет, олигохет, у некоторых насекомых (Chironomus), ракообразных, некоторых моллюсков (Planorbis corneus) и т. п.
Сравнение химического состава гемоглобина позвоночных и пигментов беспозвоночных животных показывает, что при всех различиях они обладают значительным сходством. Так (по Гриффитсу), красящее вещество крови дождевого червя содержит 53,86—53,91% С, 7,02—7,10% Н, ? N, 0,37—0,41% 5, 0,39% Fe. Состав гемоглобина у позвоночных: 51,15—54,87% С, 6,76—7,39% Н, 16,17—17,94% N, 0,39—0,85% S, 0,33—0,59% Fe. Таким образом, и там и здесь имеются одни, и те же элементы в близких соотношениях.
Сравним содержание аминокислот в гемоглобине лошади и дождевого червя. Гемоглобин лошади содержит 2,38% триптофана, 0,74% цистина, 8,31% лизина, 3,57% аргинина, 8,13% гистидина, 3,38% тирозина. Красящее вещество крови дождевого червя содержит: 4,41% триптофана, 1,41% цистина, 1,73% лизина, 10,07% аргинина, 4,68% гистидина, 3,47% тирозина. Различия значительны (чему не следует удивляться, если вспомнить разницу в систематическом положении), но общий аминокислотный состав все-таки сходен.
Эти явления показывают, что можно говорить об элементах некоторой общности физиологической организации беспозвоночных и позвоночных.
К подобным же выводам можно прийти на основании изучения функциональных особенностей целого ряда органов животных и растений. Так, например, мочевая кислота, как продукт обмена, обнаруживается у большого числа форм — от простейших (Protozoa) до хордовых. Ферменты типа трипсина и пепсина известны не только в пределах почти любой группы животного царства, но также у насекомоядных (плотоядных) растений.
Принимается, что эволюционное развитие животного мира привело к формированию двух главных стволов его: первичноротых (Protostomia) и вторичноротых (Deuterostomia). Первые включают почти всех беспозвоночных, вторые — хордовых, иглокожих и некоторые другие группы. Деление на первичноротых и вторичноротых основывается преимущественно на эмбриологических данных, т. е. на данных морфологии.
Работы целого ряда авторов показали, что деление на два больших ствола животного царства, установленное на основании эмбриологических данных, может быть обосновано и физиологической характеристикой. Так, в мышцах хордовых имеется особое органическое соединение — фосфате н. При раздражении мышцы фосфаген распадается на креатин и фосфорную кислоту. Фосфаген, или креатино-фосфат был найден в мышцах всех хордовых. Далее, в мышцах первичноротых (большинство беспозвоночных) было обнаружено вещество, близкое к фосфагену, но в нем на месте креатина был найден аргинин. Следовательно, вместо креатинофосфата, у первичноротых имеется аргининофосфат.
Эти факты биохимического родства и различий могут быть объяснены только тем, что основные типы беспозвоночных действительно образуют одну главную ветвь эволюции, а хордовые (и некоторые другие группы, в том числе иглокожие) — другую.
Нужно указать еще на один интересный, с эволюционной точки зрения, факт. В мышцах некоторых групп организмов, а именно у части иглокожих, у кишечнодышащих, а равно у асцидий обнаружены как аргинино-, так и креатинофосфаты. Это обстоятельство указывает на отсутствие резких границ между обоими главными дивергентными стволами животных и подчеркивают факт монофилетического происхождения животного царства.
Аналогичные выводы могут быть сделаны по отношению к другим данным физиологии. Значительный интерес представляют, в частности, преципитиновые реакции.
Уже давно было показано, что инородные белковые тела (сыворотки), введенные в кровь животных, вызывают образование в ней особых антител. Если, например, ввести сыворотку угря в кровь кролика, собаки и змеи (Чистович, 1899), то сыворотки крови этих животных дают с сывороткой крови угря специфический осадок, тогда как с сыворотками рыб осадка не получалось. Было проделано большое число экспериментов, приведших к аналогичным результатам. Антитела, вырабатывающиеся в крови при введении в нее чужеродных белков, получили название преципитинов.
Если ввести сыворотку человека в кровь кролика, то в ней вырабатывается преципитин, который, дает осадок с кровью человека и человекообразных обезьян, однако не дает его при введении в кровь любых других животных.
Из этого ясна близость крови человека и человекообразных обезьян. Совершенно также преципитин кролика, полученный введением ему крови лошади, давал осадок с кровью осла, но не давал его в крови других животных. Этим же методом была показана биохимическая близость крови лошади, осла и тапира; собак и лисиц, домашней свиньи и дикого кабана и т. п. Аналогичные данные были получены для ряда беспозвоночных животных. Введением в кровь кролика экстрактов растений была доказана биохимическая близость пластических веществ многих видов растений и была сделана попытка построить на данных преципитиновых реакций естественную систему некоторых растительных групп.
Как видим, изложенные данные показывают, что физиология дает пенные доказательства в пользу представлений о филогенетическом единстве живых форм.
Значительный интерес представляют также многочисленные факты паразитологии, некоторые из которых ниже указаны. Впервые Фурман (1908) сформулировал правило, по которому каждая естественная группа птиц характеризуется специфичной в видовом отношении гельминтофауной, паразитирующей только в пределах данной группы птиц. В свете этого правила интересно отметить сходство гельминтофауны американских и австралийских сумчатых, установленное Цшокке (1904). Очевидно, что это сходство показывает родство американских австралийских сумчатых. Гаррисон (1915) показал видовое сходство пухоедов, обитающих в перьевом покрове африканских страусов, американского нанду и австралийского казуара. Так как австралийская область, как материк, обособилась еще в эоцене, то указанное сходство фауны пухоедов прямо подтверждает родство перечисленных птиц и их монофилетическое происхождение.
Изложенный выше материал позволяет сделать следующие общие выводы:
1. Ряд фактов (см. а, б) показывает, что все живые организмы при всем многообразии их организации, обладают известными чертами сходства в клеточном строении и в общих им всем проявлениях жизни.
2. Каждая естественная группа организмов (см. в, г) характеризуется, при всем многообразии видов, входящих в ее состав, сходными закономерностями эмбрионального развития, единым планом строения и гомологичными органами, сходно расположенными в соответствии со сходной общей архитектоникой тела.
3. Ряд фактов физиологии и биохимии свидетельствует о сходстве между организмами различных естественных групп.
4. Все эти факты (1, 2, 3), вместе взятые, получают естественное объяснение в свете идеи о монофилетическом происхождении органического мира в целом и каждой естественной группы в частности. Если именно эта идея дает упомянутым фактам единственное естественное объяснение, то стало быть последние являются доказательствами исторической целостности органического мира.