Для целого ряда организмов, начиная с фага и кишечной палочки и кончая дрозофилой и шелковичным червем, проведено детальное генетическое картирование отдельных локусов, т. е. составлены генные карты.
Примером такого анализа могут служить исследования С. Бензера, выполненные на фаге Т4.
Фаг Т4 является бактериальным вирусом, паразитирующим на кишечной палочке Escherichia coli. Дикий тип фага Т4 одинаково хорошо лизирует штаммы Е. coli В, К и S, вызывая в бактериальных культурах появление мелких стерильных пятен — зон лизиса бактерий. Анализу подвергались мутанты фага, обозначаемые как rII. Они отличаются от дикого типа тем, что на штамме В образуют крупные пятна, на штамме К вовсе не способны размножаться, а на штамме S по характеру роста не отличимы от дикого типа, давая такие же мелкие стерильные пятна. Эти особенности мутантов rII делают их весьма удобными для исследования.
Всего было проанализировано более 2000 мутантов rII независимого происхождения. Для решения вопроса об аллельности любых двух мутаций был использован критерий комплементарности. Бактериальную культуру штамма К подвергали смешанной инфекции фаговыми частицами двух мутантов.
Отношение двух мутантных фаговых частиц к клетке-хозяину в смешанно инфицированной культуре можно рассматривать аналогично отношению гомологичных хромосом, несущих разные мутантные гены. Если совместное заражение двумя мутантами не приводит к лизису клеток штамма К, значит мутации у обоих фагов затрагивают одну и ту же функцию и произошли в одном и том же гене. Если же при совместном заражении восстанавливается дикий тип, т. е. произойдет лизис клеток штамма К, то это означает, что мутации комплементарны и произошли в разных генах. В результате такого испытания все проанализированные мутанты rII распределились на две большие группы: А и В. Мутации каждой группы мутантов были аллельными друг другу, но они не были аллельны мутациям другой группы.
Для установления частоты рекомбинации между любыми двумя мутантами rII последние были подвергнуты скрещиванию путем смешанной инфекции (обоими мутантными фагами) клеток Е. coli штамма В в жидкой среде. Предполагалось, что среди вновь развившихся частиц фага должны возникнуть как исходные нерекомбинантные мутантные формы, так и две рекомбинантные формы: двойные мутанты и дикий тип в равных количествах. Для проверки этого лизат переносили на культуру бактерии штамма В и на культуру штамма К, развивающихся на стандартной среде. В культуре штамма В вырастали как мутантные (нерекомбинантные и рекомбинантные), так и нормальные, рекомбинантные формы фаговых частиц. В культуре штамма К могли развиваться только рекомбинантные частицы дикого типа.
Поскольку частицы дикого типа и двойные мутанты появляются с одинаковой вероятностью (в результате одного события рекомбинации), их общее количество должно быть приблизительно одинаково. Для вычисления частоты рекомбинации нужно удвоенное число пятен на штамме К разделить на общее число пятен на штамме В. Тем самым будет определена частота рекомбинации между соответствующими мутациями rII.
Попарное испытание в скрещиваниях всех мутантов потребовало бы затраты огромного труда, поскольку количество необходимых опытов возрастало бы пропорционально квадрату числа испытываемых мутантов. Поэтому для облегчения работы по картированию двух соседних генов А и В были использованы мутанты с разной длиной нехваток — делеций.
Как известно, делеция представляет собой выпадение участка хромосомы. Поэтому естественно, что в этом участке невозможны обратные мутации. Исследование частоты обратных мутаций отдельных rII мутантов фага Т4 к дикому типу при культивировании их на штамме К показало, что ряд мутантов вообще не дает возврата к норме, т. е. не лизирует бактерии этого штамма и поэтому не образует стерильных пятен на штамме К. Кроме того, каждый из этих мутантов не обнаруживал рекомбинаций с некоторыми другими мутантами, которые между собой рекомбинацию выявляли. Это позволило предположить, что мутанты, неспособные к возвратным мутациям и не дающие рекомбинаций, обусловлены делениями.
Среди общего числа мутантов было найдено значительное количество таких стабильных мутантов, обусловленных нехватками. Локализацию этих стабильных мутантов производили путем попарных скрещиваний вышеописанным методом. Так была произведена грубая разбивка на отдельные участки локусов двух генов А и В на карте фага Т4. После этого приступили к более тонкому картированию с помощью перекрывающих друг друга делеций.
При скрещивании двух мутантов rII, один из которых обусловлен крупной делецией, между ними не будет происходить рекомбинации в том случае, если вторая мутация возникла в районе данной делении.
Было выбрано семь крупных перекрывающихся делеций, в сумме захватывающих почти целиком районы обоих генов А и В. Каждая из этих делеций имеет номер: r1272, r1241, r13, rPT1, rPB242, rА105 и r638. Вновь возникающий мутант скрещивали с каждым из выбранных мутантов, несущих одну из этих шести делеций. По отсутствию рекомбинации определяли примерную локализацию вновь возникшей мутации в соответствующем участке карты генов А и В. Когда на том или ином участке оказывалось Достаточное количество мутаций, путем взаимных скрещиваний определяли их линейное расположение относительно друг друга.
Если мутации являются следствием изменения участков молекулы ДНК фага Т4, то путем все более точной локализации мутаций можно чрезвычайно тонко картировать данный участок.
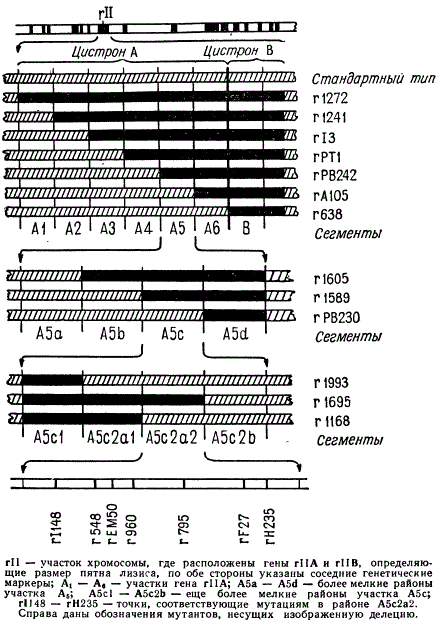
Схема тонкого картирования цистрона А фага Т4, методом перекрывающихся делений
Так, семь мутационных изменений, показанных в схеме на нижней линии, располагаются в очень коротком участке молекулы ДНК. Тщательному картированию подвергся, например, участок в районе гена А, захваченный делецией rII164.
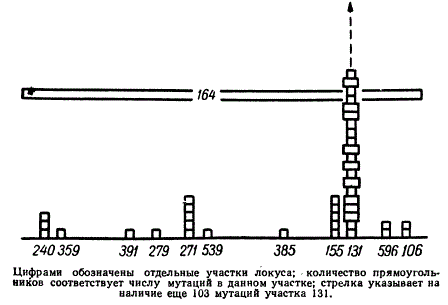
Схема распределения мутаций на участке, захваченном делецией rII164 у фага Т4
В этом участке оказались расположенными 149 мутаций. Из них 123 мутации локализовались в одной точке. Сходная картина наблюдалась и в других Участках генов А и В.
В итоге было проанализировано и нанесено на карту более 2000 мутаций локуса rII независимого происхождения.
Критерий комплементарности и испытания на рекомбинации показали, что мутации, являющиеся по своей природе точковыми изменениями или изменениями, связанными с более крупными перестройками генетического материала — делециями, распределяются в двух соседних функционально различных районах — генах А и В. Гены А и В включают более 300 точек, в которых произошло по одному или несколько мутационных изменений. Неслучайным оказался характер распределения мутаций. На карте обоих генов имелись точки, в которых мутации совершались особенно часто. Примером этого служит распределение мутаций на участке делеции rII164. Такие точки или места получили названия «горячих пятен». Такие «горячие пятна» внутри гена распределены неравномерно. Отмечено, что при этом они характерны не только для спонтанного мутирования, но и при воздействии определенными химическими агентами. Так, каждый из применявшихся аналогов пуринов или пиримидинов — 5-бромурацил, 5-бромдезоксиуръдин, 2-аминопурин, а также профлавин — вызывал свой характерный спектр распределения «горячих пятен». Специфичность действия агентов выражалась также в их влиянии на процесс обратного мутирования. Мутации, индуцированные 2-аминопурином или 5-бромурацилом, ревертируют под влиянием преимущественно этих же агентов. В то же время данные агенты не способны были вызывать реверсии к норме у спонтанных мутаций или мутаций, индуцированных профлавином. Эти факты имеют большое значение для понимания причин возникновения мутаций и самой природы генетического материала.
Исходя из признания ДНК в качестве материального носителя следственной информации, С. Бензер попытался сопоставить генетический и физический (молекулярный) масштабы карты района rII фага Т4. Известно, что ДНК фага, проникающая в бактериальную клетку, состоит примерно из 8Х104 пар нуклеотидов, взаимное расположение которых определяет генетическую информацию. Длина всей генетической карты фага Т4, измеренная в единицах рекомбинации, составляет приблизительно 800%. Отсюда нетрудно рассчитать, с какой частотой должна происходить рекомбинация между двумя соседними парами нуклеотидов. Она будет равняться 0,01% (800 : 8 : 104). Наименьшая обнаруженная С. Бензером частота рекомбинации у фага Т4 составляла 0,02%. Таким образом, на единицу рекомбинации у фага Т4 приходится не более двух пар нуклеотидов. Элементарная единица, не делимая путем рекомбинации, была названа Бензером реконом.
Мутации могут затрагивать различные по длине участки. Пока обнаружены мутационные изменения участков протяженностью не более 0,05%. Принимая во внимание столь малые размеры участков, следует признать, что единица мутации должна затрагивать изменение отдельных нуклеотидов. Наименьший участок, изменение которого может привести к возникновению мутации, Бензер назвал мутоном. Размеры мутона и рекона соответствуют одной паре нуклеотидов ДНК.
Единицу функции ген в том смысле, как мы его принимали раньше, Бензер предлагает обозначать новым термином «цистрон». Цистроном Бензер назвал участок хромосомы, в пределах которого обнаруживается цис-транс-эффект; цистрон определяет одну функцию.
Итак, тонкое картирование гена показало, что прежнее представление о гене как единице мутирования, рекомбинации и функционирования является недостаточным. Ген оказывается более сложной структурной и функциональной единицей наследственного материала, внутри которой осуществляются процессы мутирования и рекомбинации. Следует заметить, что в нашей литературе уже с 30-х годов употреблялись термины «трансген» и «базиген», соответствующие мутону и цистрону.