При изложении менделевских закономерностей наследования признаков подчеркивалось, что направление скрещивания, т. е. то, от какого пола привносятся доминантные или рецессивные признаки, не имеет значения для расщепления по данным признакам в потомстве гибрида.
Это правильно для всех случаев, когда гены находятся в аутосомах, одинаково представленных у обоих полов. В том же случае, когда гены находятся в половых хромосомах, характер наследования и расщепления обусловлен поведением половых хромосом в мейозе и их сочетанием при оплодотворении. Генетическими исследованиями установлено, что Y-хромосома гетерогаметного пола в отличие от Х-хромосомы у дрозофилы почти не содержит генов, т. е. наследственно инертна.
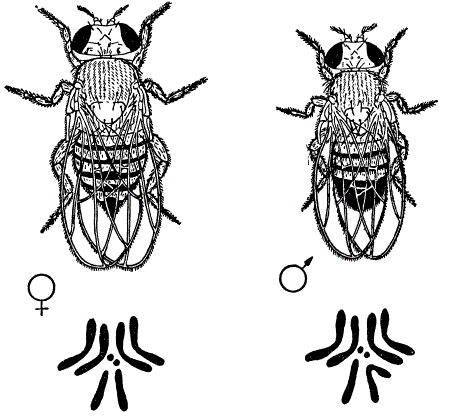
Внешний вид и хромосомные наборы (2n) самки и самца мухи-дрозофилы
Поэтому гены, находящиеся в Х-хромосоме, за некоторым исключением, не имеют своих аллельных партнеров в Y-хромосоме. Следовательно, признаки, гены которых находятся в половых хромосомах, должны наследоваться своеобразно: их распределение должно соответствовать поведению половых хромосом в мейозе. В силу этого рецессивные гены в Х-хромосоме гетерогаметного пола могут проявляться, как им не противостоят доминантные аллели в Y-хромосоме. Наследование признаков, гены которых находятся в X — и Y-хромосомах, называют наследованием, сцепленным с полом. Это явление было открыто Т. Морганом в исследованиях на дрозофиле.
Именно на этом объекте Т. Моргану с сотрудниками удалось генетическим методом показать роль хромосом в наследственности, и в частности роль половых хромосом, а также установить целый ряд общих генетических закономерностей.
Наследование при гетерогаметности мужского пола
От скрещивания белоглазых самцов дрозофилы с красноглазыми (дикого типа) самками в первом поколении все потомство (самки и самцы) оказывается красноглазым.
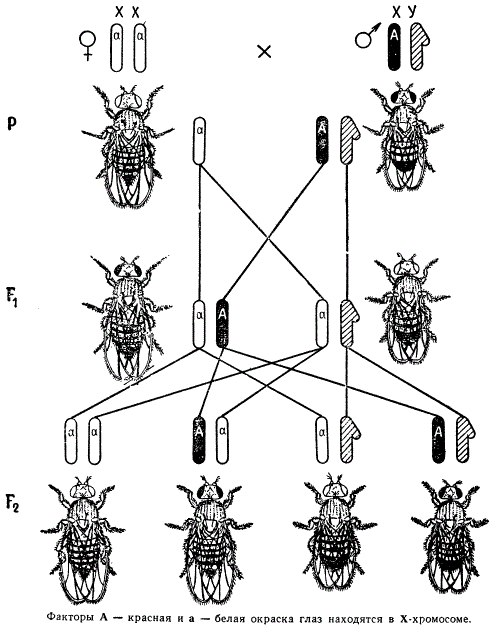
Наследование признаков, сцепленных с полом (окраска глаз) у мухи-дрозофилы
Это указывает на то, что красный цвет глаз является доминантным признаком, а белый — рецессивным. В потомстве гибридов в F2 происходит расщепление в отношении 3 красноглазых: 1 белоглазая муха, но при этом оказывается, что в норме белоглазыми бывают только самцы. На первый взгляд это кажется отступлением от менделевских закономерностей.
В случае реципрокного скрещивания, когда самка, гомозиготная по гену белых глаз, скрещивается с красноглазым самцом, расщепление наблюдается в первом же поколении в отношении белоглазых к красноглазым 1 : 1. При этом белоглазыми оказываются только самцы, а все самки — красноглазыми. В F2 появляются мухи с обоими признаками в равном отношении 1 : 1 как среди самок, так и среди самцов.
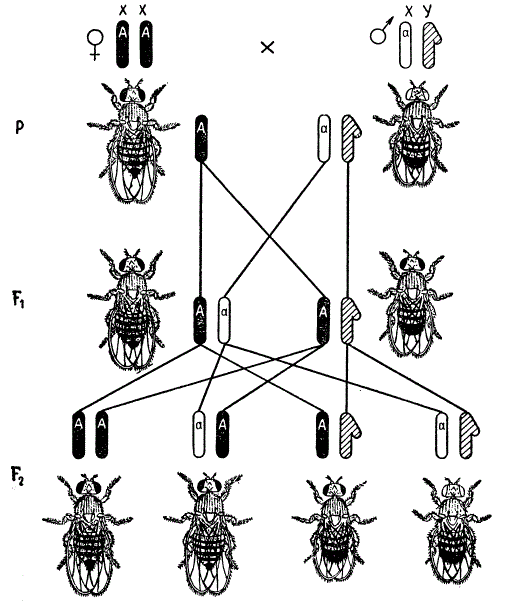
Наследование признаков, сцепленных с полом (окраска глаз) у мухи-дрозофилы. Реципрокное скрещивание
Такая закономерная связь наследования белоглазости с полом соответствует цитологической гипотезе о наследовании пола через половые хромосомы. Если предположить, что данный ген, контролирующий окраску глаз у дрозофилы, находится в X-хромосоме, тогда наследование белоглазости выглядит так, как это представлено на рисунках (на схеме не изображены аутосомы, поскольку по ним нет различия между мужским и женским организмом).
Если самка является гомозиготной по доминантному гену красной окраски глаз, находящемуся в Х-хромосоме, то этот ген вместе с половой хромосомой передается сыновьям F1, и поэтому они оказываются красноглазыми. Дочери F1 получают одну Х-хромосому с рецессивным геном белой окраски глаз от отца, а вторую Х-хромосому с доминантным геном — от матери. В силу доминирования гена красной окраски дочери оказываются также красноглазыми.
При реципрокном скрещивании дочери получают от отца одну из своих Х-хромосом, несущую доминантный ген красной окраски глаз, и одну Х-хромосому с рецессивным геном от матери, гомозиготной по гену белой окраски. Поэтому дочери F1 оказываются красноглазыми, а сыновья белоглазыми. Так как сыновья получают от матери свою единственную Х-хромосому с геном белых глаз, а от отца Y-хромосому, которая не содержит доминантной аллели красной окраски, то ген белых глаз у самца находится как бы в одной дозе. Такое состояние гена принято называть гемизиготным состоянием. Y-хромосома у дрозофилы в отношении абсолютного большинства известных признаков наследственно индифферентна. В то же время Х-хромосомы наследственно активны, и у самки каждый ген в гомологичной паре Х-хромосом представлен обоими членами аллельной пары. Для проявления рецессивного гена у самки дрозофилы необходимо, чтобы он присутствовал в обеих Х-хромосомах в гомозиготном состоянии.
Описанный тип наследования окраски глаз у дрозофилы оказался закономерным для всех организмов в отношении признаков, которые определяются генами, находящимися в Х-хромосомах. Так как половые хромосомы гомогаметного материнского организма передаются как сыновьям, так и дочерям, а единственная Х-хромосома гетерогаметного мужского пола — дочерям, то при определенном направлении скрещивания признаки, определяемые генами, находящимися в Х-хромосоме, наследуются крест-накрест, т. е. от матери — к сыновьям, а от отца — к дочерям; такое наследование признаков обычно называют наследованием крест-накрест (по английской терминологии — крисс-кросс).
Итак, наследование признаков, сцепленных с полом, полностью соответствует распределению половых хромосом в мейозе и сочетанию их при оплодотворении. На основании этого мы вправе сделать вывод о том, что гены, определяющие данные признаки, действительно находятся в половых хромосомах.
Для большей убедительности этого вывода рассмотрим еще одно скрещивание. Гены вызывают сложные физиологические изменения в процессе индивидуального развития организма. Они могут определять повышение и понижение жизнеспособности организмов вплоть до летального исхода. Такие гены могут быть рецессивными и доминантными. Рецессивные летальные гены действуют только в гомозиготном состоянии, доминантные вызывают гибель организма в гетерозиготном состоянии. Произведем скрещивание, в котором ямка дрозофилы будет гетерозиготной одновременно по двум генам — доминантному гену полосковидных глаз (В) и рецессивному гену с летальным действием (I), находящимся в Х-хромосоме: B I/B+I+.
Самец, с которым скрещивается самка указанного генотипа, в своей единственной Х-хромосоме несет рецессивный ген круглых глаз и доминантный ген нормальной жизнеспособности В+I+. Если действительно пол зависит от сочетания половых хромосом и указанные гены находятся в Х-хромосоме, то соотношение полов и расщепление по признаку формы глаз в потомстве будут находиться в точном соответствии.
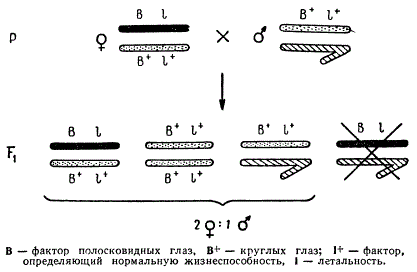
Наследование признаков, сцепленных с полом у мухи-дрозофилы
На схеме видно, что расщепление по полу оказалось не 1♀ : 1♂, а 2♀ : 1♂.
Такое отношение возникло вследствие того, что одна из Х-хромосом самки, содержащая ген В и летальный ген I, сочетаясь с Y-хромосомой, должна была определить мужской пол. Но так как в этой хромосоме находился ген с летальным действием, то такие особи на стадии личинки гибнут. Именно потому, что летальный ген находился в Х-хромосоме вместе с геном В, самцов с полосковидными глазами в потомстве не появилось. Наследование летальных генов, сцепленных с полом, является одним из неопровержимых генетических доказательств нахождения генов в хромосомах. Наследование признаков, сцепленных с полом, имеет место у многих организмов: у ряда млекопитающих, птиц, рыб, насекомых.
У человека также известен ряд примеров наследования признаков, сцепленных с полом. К ним относятся, в частности, дальтонизм (цветная слепота) и гемофилия (медленная свертываемость крови), определяемые рецессивными генами. Так как у человека гетерогаметным полом является мужской, то подобные признаки чаще проявляются у этого пола, а передатчиком таких заболеваний оказывается женский пол, который несет эти гены в гетерозиготном состоянии. Если же эти гены оказываются у эмбриона женского пола в гомозиготном состоянии, то они, как правило, вызывают смерть (в случае гемофилии).
Однако полное сцепление с полом выявляется лишь в том случае, если Y-хромосома генетически инертна. Если в Y-хромосоме имеются аллели к генам, локализованным в Х-хромосоме, то характер наследования таких признаков изменяется. Так, если мать имеет рецессивные признаки: ах ах, а отец доминантные — AXAY, то в F1 все потомки будут гетерозиготными с доминантным проявлением признака, а в F2 получится обычное расщепление на 3 особи с доминантными признаками и I — с рецессивным, причем с рецессивным признаком будут только самки. В реципрокном скрещивании в F2 с рецессивным признаком будут только самцы.
В общем виде можно сказать, что если в F2 особи с рецессивными признаками имеют тот же пол, что и родители, то такой тип наследования называется частично сцепленным с полом. Он описан для наследования некоторых признаков человека (общая цветовая слепота, кожный рак).
Гены, локализованные в Y-хромосоме и не имеющие аллелей в Х-хромосоме, наследуются отлично от других. В этом случае они наследуются только от отца к сыну. Такой характер наследования известен у человека (например, волосатые уши), у рыб и др.
Наследование при гетерогаметности женского пола
Как же осуществляется наследование, сцепленное с полом, в том случае, когда гетерогаметным полом является женский, что имеет место, например у кур, шелкопрядов, у некоторых видов рыб и других животных? У них самки несут XY, а самцы — XX хромосомы. Если верна теория сцепленного с полом наследования, то, очевидно, в этом случае все гены Х-хромосомы будут находиться в гемизиготном состоянии не у самцов, а у самок.
У кур наследуется сцепленно с полом по типу крест-накрест Целый ряд признаков: полосатое оперение плимутроков, аспидная окраска ног австралорпов и т. д. Полосатое оперение плимутроков, обусловленное особым типом распределения пигмента по перу, определяется доминантным геном, находящимся в Х-хромосоме, и проявляется в присутствии генов, обусловливающих сплошную окраску — черную, как у австралорпов, красную — типа ньюгемпширов и др. Если скрещивать полосатых кур XY с петухом XX сплошной черной окраски (породы австралорп), имеющим в гомозиготном состоянии рецессивную аллель гена полосатости, который вызывает равномерное распределение окраски, то потомство в первые же дни после вылупления можно различить по полу. Петушки, получившие доминантный ген полосатости от матери, будут иметь белое пятнышко на голове. Оперившись, они приобретут полосатую окраску. Курочки, получившие рецессивный ген сплошной окраски, оказываются одноцветными, черными.
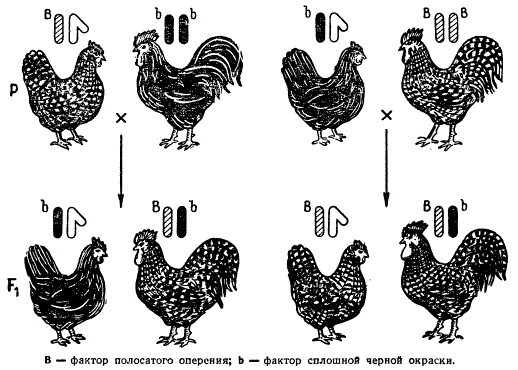
Наследование признаков, сцепленных с полом (окраска оперения) у кур
Реципрокное скрещивание курицы, имеющей сплошную черную окраску, с петухом, гомозиготным по доминантному гену полосатости, даст в F1 петухов и кур только полосатой окраски типа полосатых плимутроков.
Надо заметить, что в указанных случаях мы для простоты рассмотрели изолированное действие сцепленных с полом генов, определяющих распределение окраски, но в действительности известны гены, регулирующие выработку меланина, которые находятся как в половых хромосомах, так и в аутосомах.