Хромосомы могут утрачивать большие и маленькие участки, несущие наследственную информацию.
В метацентрической хромосоме нехватки могут возникать за счет разрывов хромосомы в различных ее точках. Соответственно числу разрывов и их месту в хромосоме образуются разные типы нехваток.
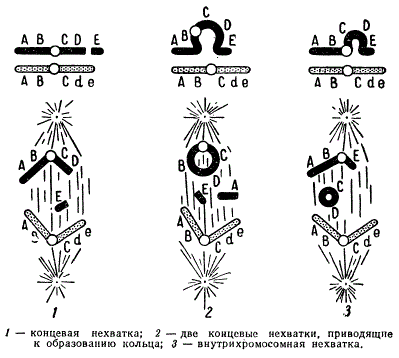
Типы нехваток хромосом
Если разрыв происходит в одном из плеч хромосомы таким образом, что один из ее концов утрачивается, то данное плечо укорачивается. Оторвавшийся фрагмент вместе с содержащимися в нем генами теряется при ближайшем делении ядра, поскольку он лишен центромеры. Такие нехватки называют терминальными (или концевыми), а также дефишенси.
Разрывы иногда происходят одновременно в двух плечах хромосомы, вследствие чего элиминируются оба ее конца. При этом открытые концы могут соединиться, образуя в митозе кольцеобразную хромосому.
Нехватки возникают также в результате двух одновременных разрывов, но не по краям, а в середине хромосомы. Места разрывов соединяются, и хромосома становится короче, при этом внутренний участок элиминируется. Если выпавший фрагмент достаточно велик, то открытые его концы могут соединиться, и в метафазе образуется ацентрическое кольцо, которое при делении ядра не ориентируется в плоскости экватора и в последующем элиминируется. Потери внутренних участков хромосом называют делециями.
Нехватки могут быть большими и малыми. К последним относят такие, которые связаны с выпадением нескольких хромомер или только части хромомеры — это микроделеции. Микроделеции можно обнаружить только при исследовании гигантских хромосом.
Мелкие нехватки обычно сохраняются в гомозиготном состоянии, давая иногда фенотипический эффект и имитируя генную мутацию. Единственный показатель, по которому их отличают от истинной мутации гена, — это отсутствие эффекта обратного мутирования.
Большие нехватки, как правило, в гомозиготном состоянии летальны, так как нарушают генный баланс. Жизнеспособными могут быть только гетерозиготы по нехваткам. В этом случае нехватки проявляются как доминантные гены. Целый ряд доминантных мутаций у дрозофилы, ранее принимавшихся за генные, оказались нехватками (Blond, Pale, Beaded, Cuvked, Plexate).
Фенотипический эффект нехватки участка хромосомы объясняют тем, что нехватка нарушает генетическую систему хромосомы, последовательность расположения генов, их взаимосвязь. Поскольку хромосомы с нехваткой при целостности центромеры сохраняют свойство репродуцировать себя в измененном виде и правильно распределяться при делении клеток, то изменение в признаке, вызванное нехваткой, наследуется. Установлено, что когда делеции и дефишенси затрагивают генетически активные районы, они почти всегда вызывают фенотипический эффект.
Следует отметить, что нехватки часто сопровождаются плейотропным фенотипическим эффектом. Нехватки вызывают понижение общей жизнеспособности и плодовитости особей.
Крупную нехватку, затрагивающую несколько дисков, можно обнаружить генетическими и цитологическими методами. Так, например, существует линия дрозофилы, где самки в одной из Х-хромосом несут нехватку части хромосомы, затрагивающую локусы вблизи гена white. Эта нехватка, названная в свое время мутацией Notch, в гетерозиготном состоянии обусловливает развитие вырезки на крыльях, т. е. мутация является доминантной.
В гомозиготном состоянии она обусловливает гибель организма, т. е. обладает рецессивным летальным действием.
Если скрестить самку дикого типа, имеющую нормальные половые хромосомы, с самцом, несущим в единственной Х-хромосоме три рецессивных гена в гемизиготном состоянии: y — определяющий желтый цвет тела, w — белые глаза и f — вильчатые щетинки,— то все особи первого поколения будут по фенотипу нормальными:
![]()
При скрещивании же самки из линии Notch, несущей в гетерозиготе нехватку локуса w, с самцом ywf в F1 половина самок будет y+W+f+/ywf дикого типа, другая половина окажется с вырезками на крыльях и белыми глазами. Очевидно, у таких самок рецессивный ген w проявился вследствие того, что доминантная аллель w+ в гомологичной хромосоме отсутствует. Другие два рецессивных гена — y и f у этих самок не проявляются, потому что их действие подавляется сохранившимися доминантными аллелями у+ и f+. Значит, выпавший участок Х-хромосомы у мух Notch относится только к району локуса w. Поэтому генотип этих мух можно представить как y+f+/ywf.
Такое явление, когда рецессивный ген проявляется в гетерозиготном состоянии, в случае выпадения доминантной аллели называют ложным доминированием, или псевдодоминированием. Рецессивный ген при этом находится в гемизиготном состоянии. Он может проявляться также у диплоидных организмов, у которых нацело отсутствует одна из гомологичных хромосом.
Можно привести также пример наследования нехватки в аутосоме у домашней мыши. У мыши известна рецессивная мутация, обусловливающая круговые «вальсирующие» движения. Если скрещивается самка, гомозиготная по этому гену w, с нормальным самцом, несущим доминантный ген w+ той же аллельной пары, то в первом поколении все потомство оказывается нормальным. Но иногда в F1 появляются «вальсирующие» особи. Когда проверили цитологически соматические клетки этих мышей, то оказалось, что одна из хромосом имела делецию. Выпадение доминантного гена w+ позволило проявиться рецессивному гену «вальсирования», находившемуся у гибрида в гемизиготном состоянии.
Однако некоторые гены, находясь в гемизиготном состоянии у самок и самцов дрозофилы, проявляются по-разному. Как было показано в опытах М. Е. Лобашева в 1935 г., при скрещивании самцов дрозофилы с геном wa (абрикосовый цвет глаз) или wco (коралловый) с самками Notch, имеющими в одной хромосоме делецию в районе локуса white, а в целой хромосоме одну из данных аллелей, в потомстве получаются два класса самок, различающихся по окраске глаз, а также по наличию или отсутствию вырезки на крыле: у самок без вырезки крыла окраска глаз сходна с окраской глаз самца, а у самок с вырезкой на крыле глаза значительно светлее, чем у самцов. Таким образом, гемизиготное состояние гена у нормального самца и самки с делецией проявляются различно: одна доза гена у самца не равна по действию одной дозе того же гена у самки. Гемизиготное состояние указанных аллелей у нормального самца и двойная доза у нормальной самки (без делеции) проявляются одинаковым образом.
Другие аллели этой серии, а именно we и wb (соответственно эозиновый и кровяной), обнаруживают иной характер действия.
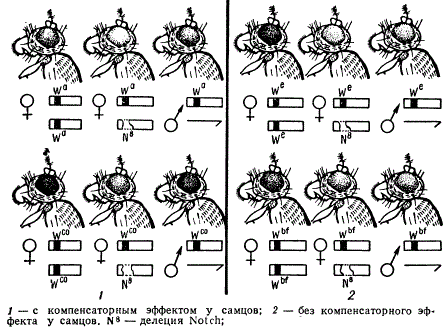
Фенотипическое проявление множественных аллелей локуса white в гомо- и гемизиготном состоянии у дрозофилы
Эти гены в гемизиготном состоянии у самок и самцов проявляются одинаково, а в двойной дозе у нормальных самок дают более темную окраску. Здесь мы не будем касаться причин, вызывающих описанные различия, так как они далеко не выяснены, однако данные результаты иллюстрируют два очень важных положения. Во-первых, выпадение Участка в одной из гомологичных хромосом дает возможность проявиться рецессивным генам в гемизиготном состоянии в другом гомологе; во-вторых, аллели в гемизиготном состоянии у самок и самцов могут проявиться не одинаково.
На основе использования гетерозиготных делеций у диплоидных организмов по схемам скрещиваний, аналогичным рассмотренной, можно обнаружить рецессивные мутации тех генов, которые противостоят нехватке в одном из двух гомологов. Так, например, у дрозофилы установлено около 130 делеций различной длины типа Notch.
На дрозофиле был разработан также специальный метод так называемых перекрывающихся делеций для локализации генов в хромосоме. Получая ряд независимых делеций разной длины в одном районе хромосомы и комбинируя их путем скрещивания с соответствующими линиями мух, можно цитологически локализовать место гена в хромосоме.

На схеме показан пример картирования генов на основе получения перекрывающихся делеций в районе white—Notch Х-хромосомы дрозофилы. Для цитологического картирования генов также использовались нехватки, возникающие в хромосомах кукурузы. Принципиально этот же метод был использован для внутригенного картирования цистронов А и В локуса rII у бактериофага Т4.
Как же осуществляется конъюгация гомологичных хромосом в мейозе в случае нехватки участка в одной из гомологичных хромосом? В мейотических хромосомах это трудно наблюдать. Лишь когда утрачивается достаточно большой участок хромосомы, такие нехватки можно обнаружить в пахитенной стадии, так как на этой стадии хромосомы выглядят в виде тонких нитей с хромомерами.
Но наиболее убедительно нехватки можно видеть в гигантских хромосомах. Гомологичные хромосомы слюнных желез дрозофилы в норме конъюгируют довольно тесно (соматическая конъюгация). При этом идентичные диски оказываются тесно прилежащими друг к другу.
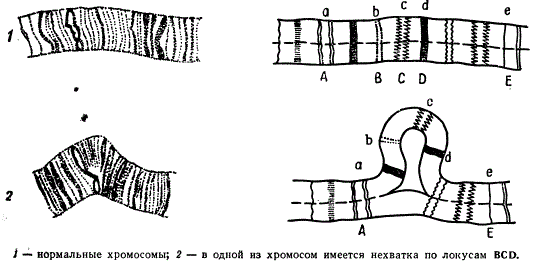
Синапсис гомологичных хромосом при наличии нехватки в одной из них
Но в случае гетерозиготного состояния по нехватке внутреннего участка одной из хромосом нормальная конъюгация в этом районе нарушается. Диски такого участка нормальной хромосомы, не имея себе партнеров в другой хромосоме, образуют петлю, в то время как все остальные гомологичные диски в обеих с хромосомах тесно прилегают друг к другу. Силы взаимного притяжения дисков в гомологичных хромосомах остаются неизвестными, но сам факт очевиден. Предполагают, что такой же тип конъюгации хромосом может иметь место и в профазе мейоза.