Можно принять за правило, что чем дальше отстоят друг от друга скрещиваемые виды и роды, тем сильнее выражена стерильность их гибридов.
Среди отдаленных скрещиваний различают две группы: 1) конгруентные скрещивания, когда родительские формы, несмотря на различие в генах, могут скрещиваться без понижения жизнеспособности, и 2) инконгруентные скрещивания, когда родительские формы имеют несоответственные хромосомы или разное число хромосом или несоответствие в плазме; у гибридов от таких скрещиваний обычно наблюдается неправильный мейоз или ненормальность в развитии и они оказываются стерильными.
Факторы, вызывающие стерильность, весьма различны:
1) несовместимость ядра и цитоплазмы и как следствие этого — нарушение митозов в процессе развития генеративных тканей;
2) действие генов, препятствующих развитию гонад и эндокринных желез у животных, а у растений — женских и мужских органов цветка;
3) генетические факторы, препятствующие конъюгации хромосом в мейозе и образованию бивалентов, следствием чего является образование гамет с несбалансированным набором хромосом.
Все эти причины обусловлены генетически, и поэтому в каждом отдельном скрещивании есть возможность установить главный фактор, определяющий бесплодие.
Атипические митозы в гибридном организме, как мы видели, могут начинаться очень рано и приводить к полной депрессии развития. Однако атипичность митозов может быть частичной и не исключает дальнейшего, даже нормального, развития гибридного организма. Атипические митозы могут проявляться в отдельных тканях, образуя химерные ткани, в том числе и в генеративной при закладке гонад, а также в процессе размножения сперматогоний и оогоний. В этом случае будет наблюдаться частичная стерильность.
У гибридов часто возникают реципрокные различия в плодовитости. Так, у некоторых гибридов семейства Bovidae межвидовые скрещивания яка и бизона с крупным рогатым скотом дают стерильных гибридных самцов и плодовитых самок. При беккроссах, по мере повышения кровности, гаметогенез восстанавливается и у мужского пола. Например, сперматогенез у гибридов первого поколения 1/2 кровного яка прекращается на стадии сперматогоний, а у 1/4 кровного яка прекращается на стадии сперматоцитов. Та же самая картина наблюдается у мула и зеброида.
Однако для полного восстановления плодовитости гибридов требуется полное замещение хромосом одного из исходных видов. Гибриды лошади и осла бесплодны в обоих полах. Сперматогенез мулов останавливается на втором делении созревания. Мулицы же иногда дают потомство как от жеребца, так и от осла. При этом беккросс на жеребца дает плодовитых гибридов, похожих на лошадь, а от скрещивания мулиц с ослом рождаются стерильные гибриды, похожие на мула. Эти факты позволяют сделать предположение, что мулицы плодовиты лишь в тех случаях, когда в оогенезе в некоторых яйцеклетках все хромосомы осла выпадают, элиминируются или попадают в направительное тельце, вследствие чего остаются лишь хромосомы лошади.
У растений отдаленная гибридизация также вызывает сходный эффект. По данным В. А. Хижняка, полноценные яйцеклетки у гибрида встречаются чаще, чем фертильная пыльца. У большинства пшенично-пырейных гибридов первого поколения в цветках пыльники не раскрываются и не содержат пыльцы.
Плодовитость отдаленных гибридов первого поколения часто зависит от выбора линий для скрещивания. Так, например, плодовитость гибридов F1 значительно выше от скрещивания Triticum durum X Agropyrom intermedium, чем от скрещивания Tr. Timopheevi X Agr. intermedium. Поскольку в обоих случаях у гибридов число хромосом одинаково (n = 21) и они различаются по плодовитости, то очевидно, что разница в плодовитости объясняется различиями геномов скрещиваемых форм.
Роды, виды и разновидности являются генетически полиморфными в отношении скрещиваемости и плодовитости гибридов.
Главной причиной, определяющей бесплодие отдаленных гибридов, является нарушение мейоза в гаметогенезе. Нарушение мейоза может быть вызвано рядом генетических факторов.
1. Различие в геномах, что ведет к нарушению равного распределения хромосом в метафазе I.
2. При общем сходстве геномов и равенстве числа хромосом скрещиваемых видов имеются различия в отдельных аллелях, а также асинаптические гены, препятствующие нормальному спариванию хромосом. Нарушение мейоза может быть вызвано не только генетическими причинами, но и неблагоприятными факторами внешней и внутренней среды. У животных особое значение приобретают гормональные факторы.
Все указанные выше причины стерильности гибридов так или иначе вызывают нарушение нормального синапсиса хромосом, что препятствует образованию бивалентов. Если биваленты не образуются, то редукционное деление нарушается, так как униваленты распределяются случайно и неравномерно. Следствием неравномерного распределения хромосом относительно полюсов в метафазе появляется образование частично или полностью неполноценных гамет. В том случае, когда один из скрещиваемых видов является автополиплоидом, а другой — диплоидом, в мейозе у гибрида могут возникать не только униваленты, но и триваленты. В случае скрещивания автополиплоидных видов могут образовываться мультиваленты, что также ведет к образованию несбалансированных
Рассмотрим нарушение нормального хода мейоза у аллополипплоидных гибридов. Каждый вид растений, произошедший путем автополиплоидии, обязательно несет несколько разных геномов, в отличие от автополиплоида, который несет сходные геномы. Чем меньше Реология геномов у скрещиваемых видов, тем ниже плодовитость гибридов. Степень плодовитости гибрида указывает на степень гомологичности геномов. Разные геномы, сочетаясь в гибридных клетках, ведут себя в мейозе по-разному. Гомологичные хромосомы сходных геномов, образуя биваленты, нормально расходятся к полюсам в анафазе I. Хромосомы разных геномов, не имеющие гомологов, не образуют бивалентов и распределяются неравномерно. Лишь иногда, когда все хромосомы отходят к одному полюсу, возможно образование нормальных гамет.
Хромосомы разных геномов имеют, как правило, морфологические отличия; часто они отличаются циклом спирализации, что проявляется в их неодновременном вступлении в метафазу.
Поскольку образование бивалентов является следствием синапсиса гомологичных хромосом, то в изучении причин бесплодия гибридов имеет значение также учет типа синапсиса отдельных хромосом и числа образующихся хиазм. По числу бивалентов, унивалентов и мультивалентов, а также по типу синапсиса (плотный и рыхлый), числу хиазм бивалентов в мейозе судят о гомологии геномов и хромосом в наборе гибридного организма.
В полиплоидном ряду рода пшениц (Triticum) диплоидная пшеница-однозернянка Tr. monococcum имеет один геном (n = 7), обычно обозначаемый А; тетраплоидные пшеницы (n = 14), например твердая пшеница (Tr. durum), имеют два разных генома А + В; гексаплоидные, например, мягкие пшеницы — три разных генома А + В + D. Таким образом, однозернянка оказывается диплоиден (АА = 2n = 14), твердая пшеница — аллотетраплоидом (АА + ВВ = 2n = 14 + 14); мягкая пшеница — аллогексаплоидом (АА + BB + DD = 2n = 14 + 14 + 14). Хромосомы генома А в мейозе, как правило, не конъюгируют с хромосомами геномов В и D.
Существует еще один вид аллотетраплоидной пшеницы Tr. Timopheevi, открытый П. М. Жуковским в 1929 г., который имел так же, как и твердая пшеница, 2n = 28 хромосом. Скрещивание этой пшеницы с твердой пшеницей осуществляется с трудом, у гибридов сильно нарушен мейоз. В случае сходства геномов в мейозе должно было бы образовываться 14 бивалентов, но фактически наблюдается 7 бивалентов и 14 унивалентов.
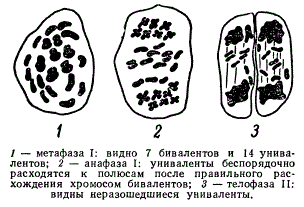
Мейоз гибрида F, Triticum durum и Тг. Timopheevi
7 хромосом у каждого из этих видов относятся к одному геному (АА), а 14 хромосом — к разным геномам. В случае скрещиваний Tr. Timopheevi (2n = 28) с однозернянкой, имеющей геном А (n = 7), в мейозе у гибрида обнаруживается менее 7 бивалентов и более 7 унивалентов. Следовательно, у Tr. Timopheevi есть геном А, сходный с таковым у однозернянки. Этот же геном выявится по числу бивалентов и у твердой 28-хромосомной пшеницы, которой геном Tr. Timopheevi, негомологичный также геному В твердой пшеницы, был обозначен G. Следовательно, у Tr. Timopheevi есть два генома АА + GG. Как было установлено, род пшениц имеет 4 разных генома: А, В, D, G.
При скрещивании видов и цитологическом анализе степени конъюгации хромосом и образования бивалентов и унивалентов в мейозе удается выяснить гомологичность геномов и причины стерильности гибридов, вызванные несовместимостью геномов. Такой цитогенетический анализ геномного состава, по предложению Г. Кихары, с 1924 г. стали называть геномным анализом.