К различным типам полового размножения можно отнести партеногенетическое, гиногенетическое и андрогенетическое размножение животных и растений.
Перечисленные типы полового размножения возникли вследствие полной или частичной утраты мейоза и замены его митозом в цикле полового размножения. Бесполое размножение в этом случае является вторичным.
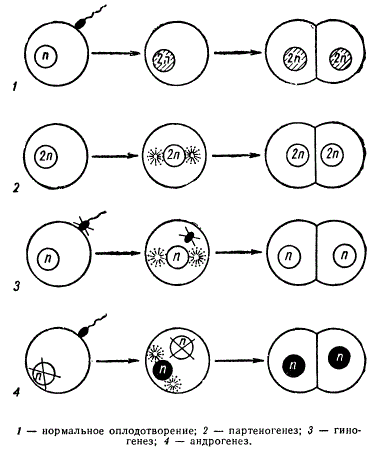
Мы уже указывали на то, что у большинства видов животных и растений при половом размножении происходит слияние двух гамет — мужской и женской. У ряда видов животных и растений имеет место девственное размножение без участия сперматозоида. Развитие зародыша из неоплодотворенной яйцеклетки называют партеногенезом.
Типы полового размножения
Партеногенез подразделяют на естественный и искусственный. При естественном партеногенезе яйцо, претерпевшее или не претерпевшее деления созревания, под влиянием внутренних или внешних причин начинает дробиться и развивается в нормальный эмбрион без какого-либо участия сперматозоида. Явление естественного партеногенеза свойственно низшим ракообразным, коловраткам, перепончатокрылым (пчелам, осам) и др. Известен он также у птиц (индейки).
Партеногенез может быть постоянным (облигатный партеногенез) или частичным (факультативный партеногенез). У одних животных из неоплодотворенных яиц могут развиваться только самки, а самцы — из оплодотворенных, у других — оба пола, у третьих — из неоплодотворенных яиц развиваются только самцы, а из оплодотворенных — самки.
Партеногенетическое размножение может перемежаться в поколениях с половым (циклический партеногенез). У низших ракообразных (дафний), а также у тлей, коловраток и других животных наблюдается смена поколений, развившихся из нормально оплодотворенных и из партеногенетических яиц. У дафний женские особи диплоидны, мужские — гаплоидны. При благоприятных внешних условиях дафнии размножаются партеногенетически. В этом случае появляется только один пол — женский, поскольку яйца не претерпевают мейотического деления. Причина этого явления станет понятна, когда будут рассмотрены генетические и цитологические механизмы определения пола. Наступление неблагоприятных внешних условий, например понижение температуры или недостаток корма, ведет к тому, что женские особи начинают откладывать гаплоидные яйца. Из этих яиц партеногенетически развиваются мужские особи. После спаривания и нормального оплодотворения восстанавливается половое поколение женских организмов с диплоидным числом хромосом. Оплодотворенные яйца в цистах могут переносить зимовку и неблагоприятные внешние условия. Сходная картина наблюдается у травяной тли и других насекомых.
Различают партеногенез соматический, или диплоидный, и генеративный, или гаплоидный. При соматическом партеногенезе яйцеклетка не претерпевает редукционного деления, или, если и претерпевает, то два гаплоидных ядра, сливаясь вместе, восстанавливают диплоидный набор хромосом (автокариогамия); таким образом, в клетках тканей зародыша сохраняется диплоидный набор хромосом. В некоторых случаях соматические клетки таких организмов могут иметь умноженное число хромосом за счет нерасхождения целых наборов хромосом. При генеративном партеногенезе зародыш развивается из гаплоидной яйцеклетки. Как правило, из таких яиц развиваются особи мужского пола (пчелы, червецы, клещи). Например, у медоносной пчелы трутни развиваются из неоплодотворенных гаплоидных яиц путем партеногенеза. В ходе развития половых клеток у них мейоз заменен митозом, и поэтому сперматозоиды имеют гаплоидный набор хромосом. В отличие от зачаткового пути в соме таких животных может восстанавливаться диплоидный набор хромосом.
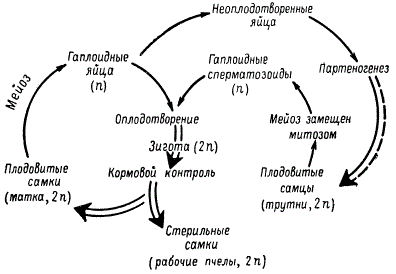
Жизненный цикл пчелы
Искусственным партеногенезом называют экспериментально вызванную активацию неоплодотворенных яиц. Честь этого открытия принадлежит русскому зоологу А. А. Тихомирову, который впервые осуществил искусственный партеногенез в 1885 г. на яйцах тутового шелкопряда. Искусственный партеногенез можно вызвать действием высокой температуры, кислот, света и других агентов. Возможность искусственного партеногенеза доказана для многих водных и наземных беспозвоночных (морские ежи, звезды, насекомые и др.) и позвоночных животных (земноводные).
При искусственном партеногенезе часто тормозится нормальное развитие зародыша. Однако, используя усовершенствованную методику обработки яиц тутового шелкопряда высокой температурой при определенной экспозиции и на определенной стадии развития, Б. Л. Астаурову удалось получить в большом количестве партеногенетические бабочки женского пола. В настоящее время искусственный партеногенез осуществлен также у лягушек и кроликов. Искусственный партеногенез получен и у растений (водоросли, грибы и высшие растения: хлебные злаки, бобовые и др.). Он стимулируется раздражением рыльца чужеродной или убитой пыльцой, а также тальком, мелом и др. В этом случае так же, как и у животных с партеногенетическим развитием, наследование осуществляется только по материнской линии.
К типу партеногенетического размножения можно отнести и гиногенетическое размножение, т. е. развитие зародыша исключительно за счет женского ядра. В отличие от партеногенеза в этом случае необходимо участие сперматозоида для стимуляции развития яйцеклетки (псевдогамия), но оплодотворения (кариогамии) в этом случае не происходит. Гиногенез обнаружен у гермафродитных круглых червей, живородящей рыбки Mollienisia formosa и у серебряного карася, встречающегося у нас на Дальнем Востоке. Гиногенез, как правило, обнаруживается у особей вида на границах его ареала как механизм, гарантирующий здесь сохранение вида.
Гиногенетическое развитие яиц можно вызвать искусственно, если перед оплодотворением сперму облучить рентгеновыми лучами, обработать химическими веществами или подвергнуть действию высокой температуры. При этом разрушается ядро сперматозоида, и оно теряет способность к кариогамии, но такой сперматозоид может активировать яйцо. Естественный и искусственный гиногенез встречается также у растений и вызывается теми же факторами, что и у животных. В случае естественного гиногенеза развивающиеся особи содержат нормальное диплоидное число хромосом. Искусственный гиногенез часто связан с гаплоидией, поэтому такие зародыши мало жизнеспособны.
Явление партеногенетического и гиногенетического размножения имеет большое значение для изучения наследственности, так как при этом потомство полностью сходно с материнским организмом. Изучение партеногенеза, как и гиногенеза, важно также и для решения ряда практических вопросов, в частности для получения особей одного определенного пола у некоторых практически важных объектов, например для разведения ценных пород рыб, а также кур.
Прямой противоположностью гиногенеза является андрогенез; при андрогенезе развитие яйца осуществляется только за счет мужских ядер, а женское ядро не принимает участия. Андрогенез может иметь место в тех случаях, когда материнское ядро почему-либо погибает до момента оплодотворения. Если в яйцеклетку попадает один сперматозоид, то развивающийся зародыш оказывается нежизнеспособным или мало жизнеспособным. Жизнеспособность андрогенных зигот нормализуется, если восстанавливается диплоидный набор хромосом. Как было уже показано, в тех случаях, когда возможна полиспермия и в яйцеклетку одновременно проникает несколько сперматозоидов от одного или от двух разных самцов (полиандрия), возможно слияние двух отцовских пронуклеусов и образование диплоидной зиготы (гетероспермное оплодотворение). Развитие андрогенных особей до взрослого состояния наблюдалось лишь у тутового шелкопряда и паразитической осы (Habrabracon hebetor). Эти особи оказываются сходными с отцовским организмом и всегда бывают только мужского пола в случае его гомогаметичности. Андрогенетическое размножение, как исключение, установлено и у растений (табак, кукуруза и др.). Болгарский генетик Д. Костов в опытах с табаком получил растение с гаплоидным набором хромосом и целиком похожее на отцовскую форму; следовательно, ядро материнского растения не принимало участия в оплодотворении, и развитие растения началось из клетки, содержащей только ядро спермия. Последующие исследования, в частности опыты Е. Н. Герасимовой с родом Crepis, подтвердили возможность такого явления.
Иногда в искусственных условиях при удалении женского ядра с частью цитоплазмы оставшаяся часть яйца после проникновения в нее сперматозоида начинает дробление, которое вскоре прекращается. Такое экспериментально вызываемое начальное развитие зародыша из части цитоплазмы яйцеклетки без участия женского ядра называют мерогенезом. В некоторых случаях (у морского ежа) дробление яйца можно получить при полном отсутствии ядра. Безъядерные партеногенетические мерогоны развивались до абортивных морул или бластул, которые вскоре погибали.
Размножение растений и животных без оплодотворения называют апомиксисом с оплодотворением (кариогамией) — амфимиксисом. Некоторые исследователи (С. С. Хохлов и др.) в понятие апомиксиса вкладывают более широкое содержание — всякое бесполое размножение, включая все его виды. Термины «апомиксис» и «амфимиксис» в равной мере приложимы к растениям и животным.
Но поскольку апомиксис особенно широко распространен в растительном мире и имеет большое значение для изучения наследования, рассмотрим его особенности. Типы апомиксиса у растений чрезвычайно разнообразны, но для них до сих пор нет общепринятой классификации.
Наиболее распространенным является тип партеногенетического образования зародыша из яйцеклетки. При этом в случае диплоидного партеногенеза полностью выпадает мейоз, а в случае гаплоидного — протекает нормальный мегаспорогенез и образуется гаплоидный зародышевый мешок с редуцированным набором хромосом в ядрах. При диплоидном партеногенезе постоянство числа хромосом двойного набора поддерживается митозом, и наследование признаков эндосперма и зародыша идет только по материнской линии.
При гаплоидном партеногенезе зародыш образуется из гаплоидной яйцеклетки также без оплодотворения. Развивающиеся из такого зародыша растения стерильные, слабые и мелколистные. Такое растение может размножаться только вегетативно. В этом случае для формирования полноценных семян необходима псевдогамия — активация зародышевого мешка пыльцевой трубкой. Один спермий из трубки, достигая зародышевого мешка, разрушается, а другой — сливается с центральным ядром и участвует только в образовании ткани эндосперма (виды лапчатки — Potent ilia, малины — Rubus и др.). Наследование здесь будет происходить несколько отлично. Признаки зародыша и растения наследуются только по материнской линии, а признаки эндосперма могут проявиться как отцовские, так и материнские.
Возникновение зародыша не из яйцеклетки, а из других клеток женского гаметофита (синергид и антипод), претерпевших и не претерпевших мейоз, называют апогаметическим. Кроме этих двух типов апомиксиса, имеются и другие, но они представляют более специальный интерес, в частности для эмбриологов растений.
Все типы апомиктического размножения (исключая вегетативное) возникли, по-видимому, в результате различных упрощений нормального полового процесса. Но как бы ни были разнообразны типы апомиксиса, если выпадает мейоз, то он замещается митозом, а оплодотворение полностью исключается. При некоторых типах апомиксиса мейоз и редукция числа хромосом сохраняются в зародышевом мешке, тогда оплодотворение замещается автогамией,
т. е. развитие зародыша осуществляется за счет слияния гаплоидных ядер самого зародышевого мешка. Последнее необходимо для поддержания постоянства видового числа хромосом. При любом типе апомиксиса не может нарушаться принцип постоянства числа хромосом вида, так как иначе число хромосом в клетках с каждым поколением будет либо увеличиваться, либо уменьшаться. Знание типа апомиксиса совершенно необходимо для изучения генетических явлений у особей, размножающихся этим способом. Примером этого может служить эпизод, имевший место в опытах Г. Менделя.
Мендель, желая выяснить универсальность открытых им закономерностей наследования на горохе, провел многолетние опыты с одним из видов рода ястребинки (Hieracium). Но у этого растения широко распространено апомиктическое размножение. Скрещивая разные расы ястребинки, Мендель не получил подтверждения желаемых результатов именно вследствие отсутствия у этого объекта процесса оплодотворения. Это обстоятельство явилось одной из причин, помешавших Менделю распространить открытые им законы на другие организмы.
Естественный партеногенез и апомиксис играют важную роль в эволюции как специальные механизмы, обеспечивающие наследственное разнообразие.
В настоящее время разрабатываются генетические приемы с использованием апомиксиса в практических целях для закрепления гетерозиса, поддержания ценных мутаций и др.