В рассмотренных до сих пор типах взаимодействия генов мы касались альтернативных, т. е. качественно различающихся, признаков.
Такие же свойства организмов, как темп роста, живой вес животного, яйценоскость кур, количество молока и его жирность у скота, шерстность овец, количество белка в эндосперме зерна кукурузы и пшеницы, содержание витаминов в растениях, скорость протекания биохимических реакций, свойства нервной деятельности животных и т. п., нельзя разложить на четкие фенотипические классы, их необходимо измерять, взвешивать, подсчитывать, т. е. оценивать в Количественном выражении. Подобные признаки чаще всего называют количественными, или мерными, признаками. Каким же образом они могут наследоваться?
Рассматривая действие гена, определяющего количество витамина А в эндосперме кукурузы, мы видели, что чем больше доза доминантного гена (YYY), тем выше активность витамина А. Допустим, что количественные признаки, образующие по своему проявлению непрерывный вариационный ряд, определяются взаимодействием многих доминантных генов, действующих на один и тот же признак или свойство. В таком случае количественно варьирующий признак у разных особей одного и того же поколения будет определяться разным числом доминантных генов в генотипе.
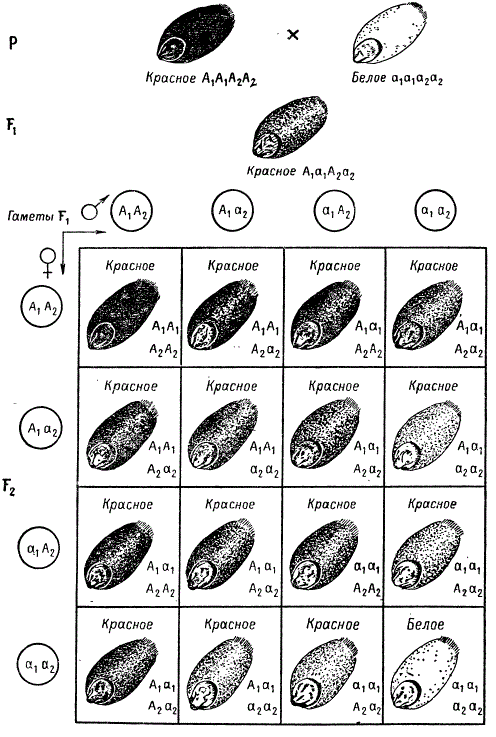
Изучение наследования полимерных признаков было начато в первом десятилетии нашего столетия. Так, при скрещивании рас пшениц с красными и белыми (неокрашенными) зернами шведский генетик Г. Нильсон-Эле в 1908 г. обнаружил в F2 обычное моногибридное расщепление в отношении 3:1. Однако при скрещивании некоторых линий пшениц, различающихся по таким же признакам, в F2 наблюдается расщепление в отношении 15/16 окрашенных и 1/16 белых, Окраска зерен из первой группы варьирует от темнокрасных до бледно-красных. Генетический анализ растений пшеницы в F3 из семян F2 разных окрасок показал, что растения, выращенные из белых зерен и из зерен с наиболее темной, (красной) окраской, в дальнейшем не дают расщепления. Из зерен с окраской промежуточного типа развились растения, давшие в последующих поколениях расщепление по окраске зерна.
Наследование окраски зерна у Triticum при взаимодействии двух пар генов (полимерия)
Анализ характера расщепления позволил установить, что в данном случае красную окраску зерен определяют две доминантные аллели двух разных генов, а сочетание их рецессивных аллелей в гомозиготном состоянии определяет отсутствие окраски. Интенсивность окраски зерен зависит от числа доминантных генов, присутствующих в генотипе.
Гены такого типа были названы полимерными, и, поскольку однозначно влияют на один и тот же признак, было принято обозначать их одной латинской буквой с указанием индекса для разных членов: А1, А2, А3 и т. д.
Следовательно, исходные родительские формы, давшие расщепление в F2 15 : 1, имели генотипы А1А1А2А2 и а1а1а2а2. Гибрид F1 обладал генотипом А1а1А2а2, а в F2 развились зерна с разным числом доминантных генов. Наличие всех четырех доминантных аллелей генов окраски А1А1А2А2 1/16 растений определяет самую интенсивную окраску зерна; 4/16 всех зерен F2 имели три доминантные аллели (типа А1А1А2а2), 6/16 — две (типа А1а1А2а2), 4/16 — одну (типа А1а1а2а2); все эти генотипы определяли различные промежуточные окраски, переходные между интенсивно красной и белой. Гомозиготной по обоим рецессивным генам (а1а1а2а2) являлась 1/16 всех зерен, и эти зерна оказались неокрашенными. Нетрудно заметить, что частоты пяти перечисленных генотипических классов при расщеплении в F2 распределяются в ряд: 1+4+6 + 4+1 = 16, который отображает изменчивость признака окраски зерна пшеницы в зависимости от числа доминантных аллелей в генотипе. Точно такой же тип наследования известен для некоторых видов окраски зерен кукурузы.
При накоплении доминантных полимерных генов их действие суммируется. Эти гены имеют кумулятивный эффект, поэтому взаимодействие такого типа называют кумулятивной полимерией.
Очевидно, что если у гибрида F1 число таких генов в гетерозиготном состоянии оказывается не два, а три А1а1А2а2А3а3 или более, то число комбинаций генотипов в F2 увеличивается. Этот ряд генотипов можно представить в виде биноминальной кривой изменчивости данного признака.
В опыте Г. Нильсона-Эле тригибридное расщепление в F2 по генам окраски зерен пшеницы давало соотношение: 63 красных и 1 неокрашенное. В F2 наблюдались все переходы от интенсивной окраски зерен с генотипом А1А1А2А2А3А3 до полного ее отсутствия у a1a1a2a2a3a3. При этом частоты генотипов с разным количеством Доминантных генов распределились в следующий ряд: 1 + 6 + 15 + 20 + 15 + 6 + 1 = 64. Приведены кривые распределения частот генотипов с разным числом доминантных генов Кумулятивного действия при независимом их сочетании в моно-, ди- и тригибридном скрещиваниях. Из этого сопоставления видно, что чем большее число доминантных генов определяет данный признак, тем больше амплитуда изменчивости.
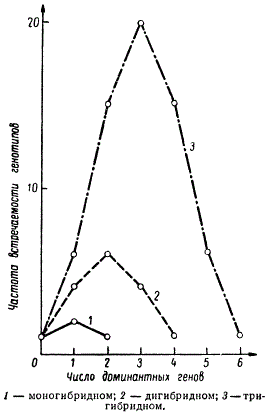
Кривые распределения частот генотипов в F2 в случае кумулятивной полимерии при скрещивании
По типу полимерных генов наследуется пигментация кожи у человека. Например, у супружеской пары негра и белой женщины урождаются дети с промежуточным цветом кожи (мулаты). У супружеской пары мулатов рождаются дети по цвету кожи всех типов окраски от черной до белой, что определяется комбинацией двух пар аллелей полимерных генов.
Изучение полимерных генов имеет не только теоретический, но и большой практический интерес. Установлено, что многие хозяйственно ценные признаки у животных и растений, такие, как молочность скота, яйценоскость кур, вес и рост животного, длина колоса, длина початка кукурузы, содержание сахара в корнеплодах свеклы, плодовитость и скороспелость животных, длина вегетационного периода у растений и многие другие наследуются по типу полимерии.
Изучая наследование перечисленных выше признаков, мы не наблюдаем в F2 расщепления на определенные, легко отличимые фенотипические классы, как это имеет место в случае альтернативно наследующихся признаков: красная или белая окраска цветка, гладкая или морщинистая форма семян у гороха и т. д. Полимерные признаки необходимо измерять. Поэтому в отличие от альтернативно наследующихся, так называемых качественных, признаков генетики различают количественные признаки. При наследовании количественного признака потомство гибрида образует непрерывный вариационный ряд по фенотипическому проявлению данного признака.
В принципе деление признаков на количественные и качественные условно. Как те, так и другие признаки можно и должно измерять при изучении их наследования, поскольку без количественной оценки любого явления природы не может быть объективного анализа. Изменчивость количественного признака в отличие от альтернативного оценивается амплитудой его варьирования.
Сама амплитуда варьирования признака наследственно определена и имеет приспособительное значение в индивидуальном развитии.
В качестве примера к сказанному приведем опыт Е. Иста по скрещиванию двух форм кукурузы — длиннопочатковой и короткопочатковой.
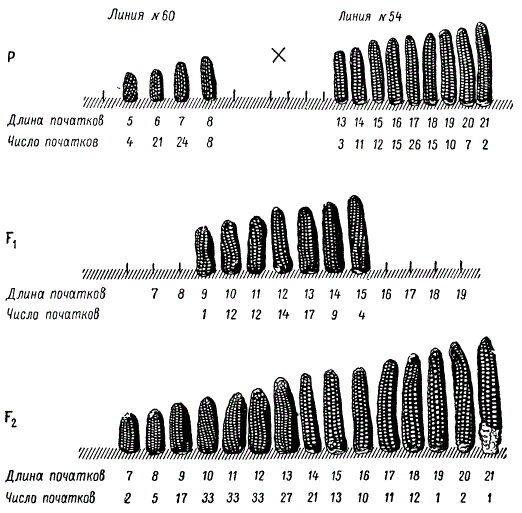
Наследование и изменчивость длины початков (в см) у Zea mays в F1 и F2
Початки по их длине у исходных линий кукурузы № 60 (короткопочатковая) и № 54 (длиннопочатковая), а также у гибридов первого и второго поколений распределяются с определенной закономерностью. Нетрудно заметить, что эти две линии сильно различаются между собой, но в пределах каждой из них длина початков колеблется незначительно. Это указывает на то, что они наследственно сравнительно однородны. Захождения в размерах початков у родительских форм нет. У гибридных растений длина початков оказывается промежуточной, с небольшой изменчивостью ряда. При самоопылении растений F1 в следующем поколении (F2) размах изменчивости по длине початков значительно увеличивается. Если вычертить кривую распределения классов по длине початков, отложив на Абсциссе размер початков, а на ординате — их количество, она оказывается сходной с кривой распределения полимерных доминантных генов. Следовательно, непрерывный ряд изменений по длине початка кукурузы можно представить как ряд генотипов при тригибридном скрещивании с различным числом доминантных генов, обусловливающих данный количественный признак.
Тот факт, что при небольшом числе исследованных растений второго поколения у некоторых из них воспроизводится длина початков, свойственная родительским формам, может указывать на участие небольшого числа полимерных генов в определении длины початка у скрещиваемых форм. Такое предположение вытекает из известной нам формулы 4n, определяющей число возможных комбинаций гамет, образующих зиготы в F2, в зависимости от числа пар генов, по которым различались исходные родительские формы. Появление в F2 растений, сходных с родительскими формами, при объеме выборки, равном 221 растению, указывает на то, что число независимо наследующихся генов, определяющих длину початка, не должно превышать трех (43 = 64) или четырех (44 = 256).
Приведенные примеры анализа наследования количественных признаков иллюстрируют лишь один из возможных путей изучения сложных и колеблющихся в своем проявлении признаков. Большая изменчивость признака, прежде всего, указывает на его сложную генетическую обусловленность, и, напротив, меньшая изменчивость признака — на меньшее число факторов, его определяющих.
Для изучения наследования количественных признаков и их изменчивости необходимо применение статистических методов обработки и оценки измерений. Мы не будем касаться этих методов, так как их можно найти в ряде учебников и пособий по вариационной статистике.
Данные, полученные в результате изучения наследования количественных признаков, явились одним из первых доказательств того, что признаки организма обусловлены многими генами. Анализ наследования и действия этих генов чрезвычайно сложен, потому что, во-первых, число полимерных генов, могущих определять развитие даже одного количественного признака, изменчиво, во-вторых, сила действия и значение каждого из этих генов могут быть специфичны, в-третьих, каждый из этих генов может иметь разную степень доминирования.
Несмотря на сложность изучения наследования и действия множественных генов, необходимо признать, что теория множественных генов оказывается пока наилучшим объяснением закономерностей для наследования количественных признаков. Она раскрывает возможности для успешной работы селекционера по созданию сортов растений и пород животных.
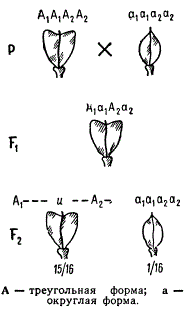
Наследование формы стручка у Capsella bursa pastoris при взаимодействии двух пар генов (полимерия)
Полимерные гены с однозначным действием могут наследственно определять и качественные, т. е. альтернативные, признаки. Примером такого действия полимерных генов может служить наследование формы плода (стручка) у пастушьей сумки. У этого вида обычно встречаются растения с треугольной и очень редко — с яйцевидной формой плода. От скрещивания этих рас в F1 появляются растения, которые имеют плоды треугольной формы. Во втором поколении происходит расщепление по фенотипу в отношении 15/16 с треугольными стручками и 1/16 в с яйцевидными, т. е. наблюдаются два фенотипических класса.
Предположим, что раса пастушьей сумки (Capsella bursa pastoris) с треугольными стручками гомозиготна по двум парам однозначных доминантных генов (генотип A1A1A2А2), а раса с яйцевидными стручками имеет генотип a1a1a2a2. Сочетание гамет при оплодотворении дает гибриды с генотипом А1а1А2а2. Доминантные аллели каждого из двух генов действуют качественно однозначно, т. е. определяют треугольную форму стручка. Поэтому генотипы A1—A2—(9/16), A1—a2a2(3/16) и a1a1A2—(3/16) будут определять треугольную форму стручков, а яйцевидная форма стручка будет обусловливаться двойным рецессивом а1а1а2а2, и из 16 комбинаций ожидается один такой генотип.
Таким же образом осуществляется наследование оперенности голени у цыплят. При скрещивании кур, гомозиготных по двум различным рецессивным аллелям и имеющим неоперенную голень, с формами, гомозиготными по доминантным аллелям генов, определяющих оперенность голени, в F1 все цыплята имеют оперенную голень. В F2 по этому признаку наблюдается расщепление в отношении 15 : 1.
При тригибридном скрещивании расщепление по таким генам будет соответствовать отношению 63 : 1 и т. д. Такого рода доминантные однозначные гены иногда называют генами с некумулятивным действием, а явление подобного взаимодействия — некумулятивной полимерией. Рецессивные гены, обусловливающие мутантный фенотип при одновременном их присутствии в генотипе в гомозиготном состоянии, называют дупликатными генами.
Как мы видим, в случае некумулятивной полимерии наличие в генотипе разного количества доминантных полимерных генов однозначного действия не изменяет выраженности признака. Достаточно одной доминантной аллели любого из генов, чтобы вызвать развитие признака.
Некоторые авторы рассматривают явление взаимодействия однозначных генов как пример взаимного двойного (дупликатного) доминантного эпистаза: А— > B— и В— > А—. Возможно, что для ряда случаев данное объяснение вполне оправданно. Однако в отношении большинства случаев это объяснение взаимодействия полимерных генов, вероятно, неприменимо.
Происхождение полимерных генов в процессе эволюции может быть объяснено следующим образом. При случайном скрещивании двух близких форм, имеющих одинаковые доминантные гены с одним и тем же действием A1A1 и А2А2, у гибрида могло произойти удвоение набора хромосом (по типу амфидиплоидии), в результате чего новый вид будет нести оба доминантных гена: A1/A1 A2/A2.
Возможны и другие механизмы возникновения полимерных генов путем различных хромосомных перестроек. Дупликатные рецессивные гены могут возникнуть из дупликатных доминантных генов за счет мутаций: A1—→ a1 и A2—→ a2.
Однозначное действие множественных генов оказывается довольно широко распространенным явлением, оно описано для многих растений: кукурузы, пшеницы, табака, хлопчатника и др.
Мы разобрали три типа взаимодействия генов: комплементарное, эпистатическое и полимерное. Все они видоизменяют классическую формулу расщепления по фенотипу, установленную Менделем для дигибридного скрещивания 9:3:3: 1. Приведены некоторые типы расщепления по фенотипу для дигибридного скрещивания, при этом все они показаны с точки зрения доминантного и рецессивного эпистаза.
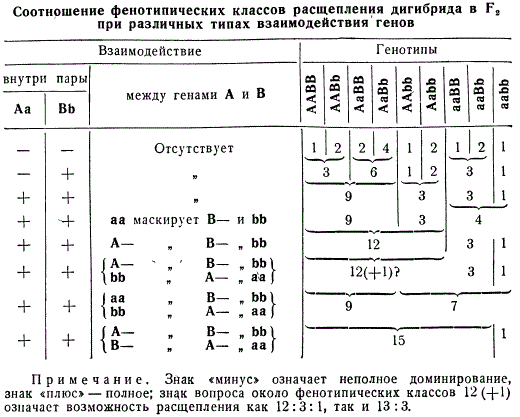
Таким образом, все приведенные типы расщепления по фенотипу столь же закономерны, как 9:3:3:1; они являются не следствием нарушения генетического механизма расщепления, а результатом взаимодействия генов между собой в индивидуальном развитии. Именно знание генетического механизма позволило выяснить эти сложные случаи наследования.
Мы иллюстрировали типы взаимодействия генов на примере дигибридного расщепления. Подобные же типы взаимодействия могут иметь место и при любом числе аллельных пар генов, наследующихся независимо друг от друга. В этом случае расщепление по фенотипу будет еще более сложным, и его можно выявить лишь при учете большого числа особей в потомстве. Так, для тетра- и пентагибридных скрещиваний необходимо иметь несколько тысяч потомков, чтобы обнаружить все классы расщепления.