Вакуоли есть почти во всех взрослых живых растительных клетках.
Они представляют собой полости внутри протопласта, заполненные обычно водянистым содержимым — клеточным соком. Так как вакуоли образуются в результате обмена веществ протопласта, то их форма, размеры и состояние определяются состоянием самого протопласта. В очень молодых, эмбриональных клетках протопласт обычно занимает весь объем клетки и вакуолей нет. В более взрослых клетках наблюдаются многочисленные очень мелкие (2—10 мк) вакуоли, равномерно распределенные в цитоплазме. Ядро обычно лежит в центре клетки. При рассматривании в световой микроскоп эти мелкие вакуоли имеют вид отдельных изолированных зерен или тонких изогнутых нитей, по форме напоминающих митохондрии. Благодаря своей многочисленности они придают цитоплазме пенистый вид. Содержимое их отличается довольно высокой плотностью и вязкостью и представляет собой гидрогель, образованный, по-видимому, гидрофильными белками. При постепенном переходе клетки во взрослое состояние, что выражается прежде всего в ее росте, объем клетки сильно увеличивается, тогда как объем цитоплазмы увеличивается незначительно. Этот процесс, называемый процессом растяжения клетки, связан с накоплением большого количества воды, поглощаемой клеткой извне, и ростом оболочки. Цитоплазма, поглощая воду, выделяет ее затем в вакуоли вместе с продуктами своей жизнедеятельности — продуктами обмена, в виде клеточного сока. При этом мелкие вакуоли растут, содержимое их разжижается, они сливаются друг с другом и число их уменьшается. Отдельные вакуоли часто принимают неправильную форму, изменяемую движением цитоплазмы. Наконец, во взрослой клетке, достигшей своего окончательного размера, все вакуоли сливаются в одну центральную вакуолю, а протопласт оттесняется к оболочке, облекая вакуолю в виде тонкого постенного слоя. Как показали электронномикроскопические исследования, толщина этого постенного слоя цитоплазмы может быть значительно меньше толщины первичной оболочки и митохондрий. Это наблюдается, например, в клетках основной паренхимы стебля, толщина слоя цитоплазмы которых находится на пределе разрешающей способности светового микроскопа (около 0,2 мк). В этих клетках плазмалемма и тонопласт до такой степени сближаются друг с другом, что клеточные органоиды (митохондрии и пластиды), зажатые между ними, изменяют свою форму.
В постенном слое цитоплазмы располагаются ядро и другие органоиды клетки. Иногда ядро занимает центр клетки, окружающая его цитоплазма соединяется с постенной цитоплазмой тяжами, проходящими через полость вакуоли. Кроме цитоплазмы, вакуолеподобные образования могут возникать при особых условиях и в других органоидах, например, в пластидах и в ядре.
Присутствие одной крупной вакуоли, заполненной клеточным соком, является характерной особенностью дифференцированной (взрослой) растительной клетки, которая остается живой к моменту зрелости. Объем такой вакуоли обычно значительно больше объема всех других клеточных компонентов, вместе взятых, и часто почти равен объему всей клетки. Так, вакуоли клеток сочных органов растений нередко занимают свыше 90% объема клетки.
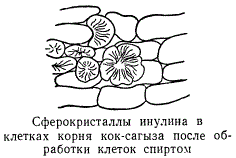
Процесс вакуолизации может быть обратимым. Так, иногда взрослые клетки опять переходят в эмбриональное состояние, приобретая способность делиться. При этом объем протопласта увеличивается, а объем клеточного сока уменьшается, вместо одной крупной вакуоли опять возникают многочисленные мелкие вакуоли, имеющие вид зернышек или коротких палочек. Содержимое вакуолей — клеточный сок — представляет собой очень маловязкую жидкость и является весьма слабым водным раствором различных веществ, синтезированных и выделенных протопластом. Таким образом, основной компонент клеточного сока — вода. В ней аккумулируются многочисленные соединения, минеральные и органические, которые находятся в состоянии истинного или коллоидного раствора и реже — в виде твердых включений. Вязкость клеточного сока связана с присутствием коллоидов, которые иногда при обезвоживании клетки могут придавать ему состояние настоящего геля. Реакция клеточного сока обычно слабокислая или нейтральная, реже щелочная. Среди веществ клеточного сока преобладают соли, органические кислоты и растворимые углеводы. Соли минеральных и органических кислот — нитраты, сульфаты, соли лимонной, щавелевой, янтарной кислот — играют наибольшую роль в создании осмотического давления клетки. Роль органических кислот клеточного сока до конца еще не выяснена. До недавнего времени их рассматривали как отбросы, конечные продукты обмена веществ протопласта. Однако появились данные, показывающие, что при определенных условиях органические кислоты могут вновь использоваться протопластом. Им приписывается также роль защиты клетки от нападения паразитов. Из растворимых углеводов в клеточном соке наиболее распространены сахароза, глюкоза (виноградный сахар) и фруктоза. Они играют роль запасных энергетических веществ и служат важнейшим питательным материалом клетки. Накапливающаяся в большом количестве в клеточном соке корнеплодов сахарной свеклы и сердцевины сахарного тростника сахароза имеет большое народнохозяйственное значение, так как служит основным источником получения сахара. Глюкоза и фруктоза, как показывает их название, распространены в плодах. В клубнях георгины, земляной груши, в корнях одуванчика и других растений семейства сложноцветных клеточный сок содержит близкий к крахмалу углевод инулин, отличающийся от крахмала растворимостью в воде. При действии спирта инулин кристаллизуется, образуя так называемые сферокристаллы.
 Для некоторых растительных групп (семейства кактусовых, толстянковых, орхидных) характерно накопление в клеточном соке слизистых веществ, также являющихся углеводами. Очень часто в состав клеточного сока входят глюкозиды (миндаль, наперстянка) и алкалоиды (мак, кофе, чай). Первые представляют собой соединения глюкозы со спиртами, альдегидами и другими веществами, не содержащими азот, а вторые — азотистые вещества сложного состава. Роль их в обмене веществ не выяснена. Они имеют горький вкус и в определенных количествах ядовиты для животных, предохраняя таким образом растение от поедания. В то же время многие из них представляют собой ценные лекарства, например атропин, — у белладонны, морфин и кодеин — у мака, хинин — у хинного дерева.
Для некоторых растительных групп (семейства кактусовых, толстянковых, орхидных) характерно накопление в клеточном соке слизистых веществ, также являющихся углеводами. Очень часто в состав клеточного сока входят глюкозиды (миндаль, наперстянка) и алкалоиды (мак, кофе, чай). Первые представляют собой соединения глюкозы со спиртами, альдегидами и другими веществами, не содержащими азот, а вторые — азотистые вещества сложного состава. Роль их в обмене веществ не выяснена. Они имеют горький вкус и в определенных количествах ядовиты для животных, предохраняя таким образом растение от поедания. В то же время многие из них представляют собой ценные лекарства, например атропин, — у белладонны, морфин и кодеин — у мака, хинин — у хинного дерева.
В клеточном соке очень часто встречаются дубильные вещества — танниды. Это сложные органические безазотистые соединения вяжущего вкуса, сильно преломляющие свет. Клеточный сок, содержащий танниды, отличается высокой вязкостью. Особенно богаты дубильными веществами клетки коры (дуб, ива, ель), листья чая, семена кофе. При отмирании клетки танниды окисляются, пропитывают клеточную оболочку и придают ей темно-коричневый цвет. Значение дубильных веществ в жизни самого растения выяснено недостаточно. Они обладают антисептическими свойствами и поэтому служат защитными веществами против нападения различных микроорганизмов. Техническое значение таннидов состоит в том, что с их помощью дубят кожу, после чего она становится мягкой, не ослизняющейся и не пропускает воду.
Все эти вещества, растворенные в клеточном соке, как правило, бесцветны и их выявляют лишь специальными реактивами. Поэтому клеточный сок может быть и бесцветным, и окрашенным в различные цвета, благодаря присутствию растворимых в воде пигментов. Наиболее распространенные пигменты клеточного сока — антоцианины и флавоны — относятся к группе глюкозидов. Чаще всего они сосредоточены в клеточном соке наружных слоев клеток высших растений. Антоцианины обусловливают красный цвет корнеплодов и листьев столовой свеклы, красный, пурпуровый или синий цвет лепестков многих цветков и других частей растений. Особенно часто они встречаются в клетках проростков и молодых растений, которые приобретают поэтому красноватые тона. Различие в оттенках цвета — от фиолетового до красного — связано с различной реакцией клеточного сока: если реакция кислая, то господствуют красные тона, при нейтральной реакции — фиолетовые, а при слабощелочной— синие. Присутствием антоцианов объясняется и цвет плодов вишни, сливы, винограда. Желтый цвет цветков, например, лепестков льнянки, желтой георгины связан с присутствием в клеточном соке пигментов группы флавонов.
Значение пигментов клеточного сока в обмене веществ выяснено недостаточно. Находясь в клетках лепестков и вызывая их яркую окраску, пигменты выполняют функцию привлечения насекомых-опылителей. Так как они сильно поглощают ультрафиолетовые лучи, то возможно, что молодые части растений благодаря этому защищены от вредного действия этих лучей.
Состав, концентрация и вязкость клеточного сока у разных видов растений различны и изменяются даже в одном растении от органа к органу, от ткани к ткани и от клетки к клетке. Поэтому за исключением воды не все клетки накапливают в вакуолях все перечисленные вещества. Многие из веществ клеточного сока, например, алкалоиды, глюкозиды встречаются только у некоторых групп растений, другие же вещества распространены более широко. Весьма часто в клеточном соке отдельных специализированных взрослых клеток накапливается практически только один продукт обмена веществ, но в больших количествах. Например, дубильные вещества накапливаются в особых крупных клетках — вместилищах, рассеянных в коре и древесине. В клеточном соке некоторых клеток могут накапливаться большие количества слизи, растворимых белков (слизевые и белковые вакуоли).
На состав и свойства клеточного сока большое влияние оказывают возраст клетки (и самого растения) и окружающие условия. Например, незрелые сочные плоды, обычно зеленые, кислые и часто вяжущие, по созревании меняют свою окраску и вкус (вишня, различные ягоды и др.). Это связано с тем, что по мере созревания плодов уменьшается содержание органических кислот, вызывающих кислый вкус, дубильных веществ, вызывающих вяжущий вкус, и накапливаются сахара. Накапливание антоцианинов особенно интенсивно происходит в листьях осенью при сухой, солнечной и прохладной погоде, когда желтеющие листья приобретают красивые красноватые оттенки, обусловленные накоплением антоцианинов.
Несмотря на то, что вакуоли с клеточным соком не обладают свойствами живого, тем не менее их значение в жизни клетки и растения очень разнообразно. Прежде всего вакуоля вместе с цитоплазмой выполняет функцию поглощения воды и растворов и передвижения их по растению. Поглощенная клеточным соком вода придает клетке упругое состояние (тургор). Тургор обеспечивает сохранение сочными органами определенной формы и положения в пространстве, а также сопротивление их действию механических факторов. Вакуоли служат также резервуарами запасной воды. Растворенные в клеточном соке соли, органические кислоты, углеводы и белки могут вновь использоваться в обмене веществ протопласта.
Механизм заложения вакуолей еще полностью не выяснен. Электронномикроскопические исследования показали, что во взрослой клетке вакуоли отграничены от цитоплазмы одной мембраной— тонопластом. Иногда, если в клетке несколько крупных вакуолей, то у них наблюдаются длинные трубки, вытягивающиеся в сторону цитоплазмы, причем трубки часто напоминают контуры гладкой эндоплазматической сети.
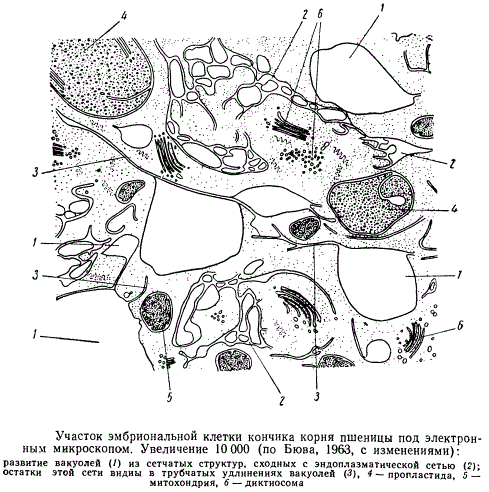
Участок эмбриональной клетки
В молодых клетках, как видно на некоторых электронограммах, обнаруживаются многочисленные местные расширения межмембранного пространства эндоплазматической сети. Эти клетки при рассматривании в световой микроскоп имеют мельчайшие вакуоли, напоминающие по форме те, которые получились на электронограммах. Это дало основание некоторым ученым выдвинуть гипотезу, согласно которой вакуоли закладываются в результате местных расширений межмембранного промежутка эндоплазматической сети. В пользу этой точки зрения свидетельствует и наличие одной мембраны вокруг вакуолей. Однако до сих пор не удалось получить электронограмм, показывающих на одном срезе непрерывность ядерной оболочки, эндоплазматической сети и вакуолей. Поэтому была развита и другая гипотеза, согласно которой в отдельных участках гиалоплазмы происходит местная гидратация (оводнение) белков без всякой связи с эндоплазматической сетью. В гиалоплазме эмбриональных клеток на электронограммах были обнаружены отдельные более светлые участки, не ограниченные сначала мембраной и содержащие остатки цитоплазмы. Эти участки и считают зачатками вакуолей. При последующем слиянии этих маленьких капелек в более взрослых клетках возникал тонопласт, и вакуоля принимала типичную форму. Возникновение трубчатых удлинений у крупных вакуолей, выступающих в гиалоплазму, по этой гипотезе, объясняется деформацией вакуолей в результате движения цитоплазмы. Ограниченные мембраной структуры с признаками вакуолей были найдены в контакте с диктиосомами или близко от них. Это послужило основанием для гипотезы, согласно которой вакуоли образуются путем разбухания межмембранного пространства наружных цистерн диктиосом. При этом мембраны диктиосомы становятся мембранами тонопласта.
Какая из этих гипотез соответствует действительности, должны показать дальнейшие исследования. Вполне возможно, что существование различных гипотез объясняется различными путями заложения вакуолей.