Широкое применение различных методов вегетативного размножения, особенно размножения черенками, а также прививок, выполняемых с разнообразными целями, побуждают исследовать те структурные процессы в каллюсах, которые определяют образование придаточных корней и почек в черенках и срастание в прививках. Можно объяснить только общие принципы соответствующих структурных процессов вследствие слабой изученности анатомической стороны методов черенкования и прививок.
На всяком срезе стебля, корня, а иногда и листа, если участки этих органов со срезами помещены в соответствующие благоприятные условия температуры и влажности, через некоторое время образуются наросты ткани беловатого или желтоватого цвета, которые по сходству их с мозолями называют мозолем, или каллюсом. В таких мозолистых, или каллюсовых наростах со временем возникают корешки и почки новых побегов. Каллюс образуется также при заживлении нанесенных растению ран и при срастании привоя с подвоем.
Начало возникновения каллюса принципиально возможно во всех растительных тканях, где в достаточном числе сохранились живые клетки. На отрезках осевых органов, стеблей или корней каллюс может образоваться из различных клеток коры, из древесины или сердцевины. Даже на разрезах таких тонких органов, как листья, при соответствующих условиях вполне может образоваться каллюс, только не столь заметный (вследствие малых размеров), как на срезах стебля и в особенности корня. На разрезах древесины каллюс образуется из тилл, заполняющих полости сосудов, и из клеток древесинной паренхимы. Но особенно легко и быстро каллюс возникает из меристематических тканей или тканей, способных быстро переходить в меристематическое состояние, в частности из камбия и перицикла.
До сих пор у самых разнообразных растений не найдено никаких особых отличий в структуре ткани каллюсов. Всюду эта ткань состоит из паренхимных клеток весьма разнообразных размеров и очертаний, расположенных как будто бы беспорядочно. Клетки каллюса вполне живые, нередко с хорошо заметными клеточными ядрами. Хотя межклетники в каллюсовой ткани развиты очень слабо и встречаются лишь изредка, однако эта ткань не меристематическая: на последнее указывают относительно толстые клеточные оболочки. Но, не имея характерных внешних признаков меристемы, ткань каллюса легко переходит в меристематическое состояние. В отдельных местах каллюсовой ткани под воздействием того или другого раздражителя возникают новые клетки. Кроме того, в больших вакуолях каллюсовых клеток, по-видимому, содержатся вещества, способные послужить источником питания для всех новообразований, возникающих в каллюсах до тех пор, пока последние не включаются в общую проводящую систему или не начнут жить самостоятельно. При образовании структуры каллюсовой ткани из нее может возникнуть любой орган растения. В смысле образовательной способности ткань каллюса весьма многогранна (тотипотентна) и при этом содержит в себе вещества, необходимые для начального развития возникающих органов.
На поверхности каллюса, хотя ткань его и однообразна, часто намечается подобие покровной ткани, состоящей из более мелких и морфологически совершенно однородных клеток, несколько похожих на таблитчатый тип эпидермиса, но без устьиц. Спустя некоторое время поверхностные клетки каллюса подсыхают, образуя подобие одного слоя пробковой ткани, или же слущиваются, заменяясь новыми слоями каллюсовой ткани.
В своем распространении ткань каллюса не ограничивается только поверхностью среза, но возникает и на некоторой глубине тканей черенка. Делению подвергаются паренхимные ткани, расположенные в коре черенка, камбий и перицикл. Раздражение, исходящее от первично образовавшейся каллюсовой ткани, с расстоянием ослабляется; чем глубже лежат ткани по оси черенка, тем энергия образования каллюсоподобных клеток слабее. В итоге массив каллюсовой ткани приобретает очертания клина, как бы вонзающегося своим острым кондом в толщу ткани черенка.
На рисунке изображена схема продольного разреза верхушки черенка шиповника с двумя клиньями каллюсовой ткани: первый клин — между первичной корой (2) и лубяными волокнами (3); второй клин более обширный, изображена только верхушка — между лубяными волокнами и древесиной центрального цилиндра (4). Такая клинообразная в продольном сечении ткань окружает кольцом весь цилиндр древесины в верхушке черенка, а также оторачивает кольцо лубяных волокон, обнаруживаясь на поверхности сначала двумя концентрическими кругами. Вскоре круги каллюсовой ткани разрастаются, сливаясь на поверхности среза черенка в одну общую кольцевую массу.
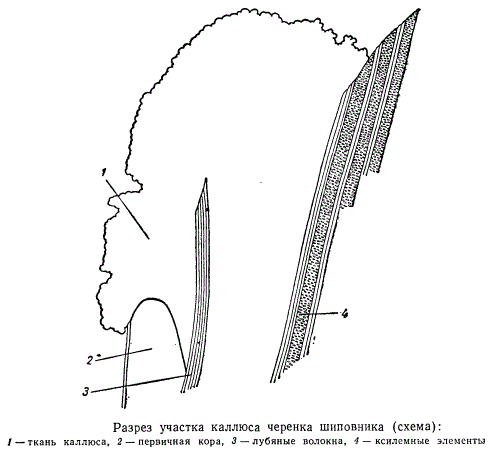
По существу срастаются друг с другом отдельные кольца каллюсовой ткани. В то же самое время в определенных участках каллюсовой ткани начинается процесс дифференциации клеток. Выше уже отмечалось, что каллюсовые клетки вполне жизнедеятельны. Особенно выделяются своей жизнеспособностью эмбриональные клетки каллюса, образовавшиеся в вышеописанных клиньях. По-видимому, эти качества зависят от того, что клетки, заполняющие клинья, произошли от камбия и перицикла и непосредственно связаны с основными образовательными тканями центрального цилиндра.
На рисунке изображена часть поперечного разреза каллюса шиповника, расположенная в толще клина. Клетки каллюсовой ткани, с первого взгляда столь беспорядочно нагроможденные в месте среза, в области клина располагаются правильными рядами, послойно размещенными между тканью первичной коры (1) и перициклическими волокнами (3).
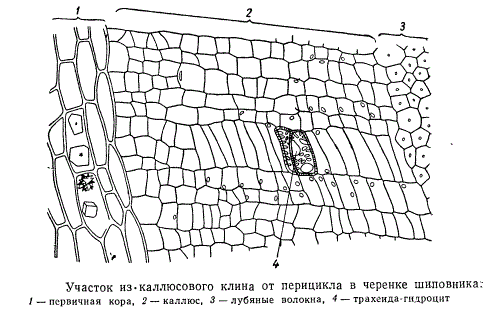
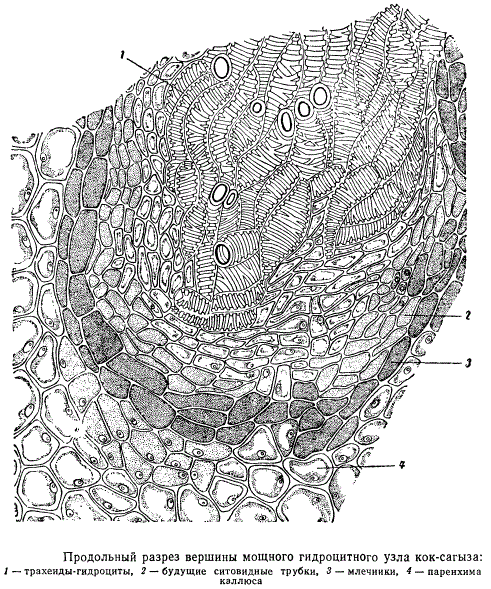
В такой ткани особенно легко возникают участки меристемы каллюса. Возможно, что меристема с определенной направленностью деятельности распространяется горизонтально от живых клеток центрального цилиндра. В итоге из возникших в отдельных местах каллюса меристематических участков начинают формироваться первые элементы сосудистой системы — трахеиды, а около них, кнаружи, т. е. в сторону первичной коры, — ситовидные трубки. На продольном разрезе верхушки черенка образовавшиеся трахеидоподобные клетки расположены цепочкой; около них цепочкой же располагаются вновь образовавшиеся ситовидные трубки. От первичной коры каллюс отделен слоем кристаллоносной паренхимы.
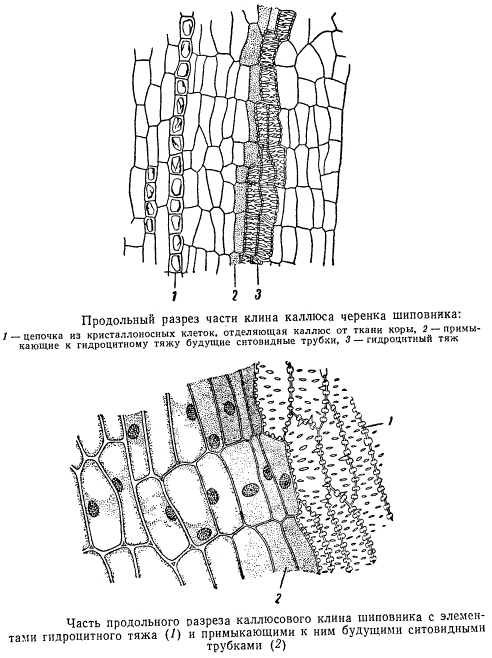
Участок тяжа, состоящего из трахеидоподобных клеток и ситовидных трубок, при несколько большем увеличении изображен на рисунке. Клетки ситовидных трубок отличаются от клеток прочей паренхимы каллюса своей вытянутостью и соответственно более мелким просветом. Трахеидоподобные клетки принадлежат к широкораспространенному типу водоносных элементов, называемых гидроцитами. Гидроциты каллюса отличаются от водоносных анатомических элементов других типов тем, что в них сравнительно долго сохраняются клеточные ядра, хотя утолщения их оболочек одревесневшие. Одревеснение клеточных оболочек отнюдь не обязательно связано с разрушением живого протопласта клеток. Например, клетки древесинной паренхимы, с определенно одревесневшими оболочками, часто вполне жизнедеятельны, имеют клеточные ядра и способны к делению. В члениках сосудов, а также у большинства типов трахеид живое содержимое клеток отмирает и разрушается.
На рисунке изображено два участка ткани, содержащей гидроциты, в которой местами прекрасно различимы клеточные ядра, а на другом рисунке — длинный гидроцит с тремя сохранившимися клеточными ядрами и участками протопласта, съежившимися, однако, при приготовлении препарата и поэтому отставшими от оболочки. От одного из клеточных ядер к периферии цитоплазмы тянутся цитоплазменные тяжи. Длинные гидроциты, как и ситовидные трубки, могут быть многоядерными.
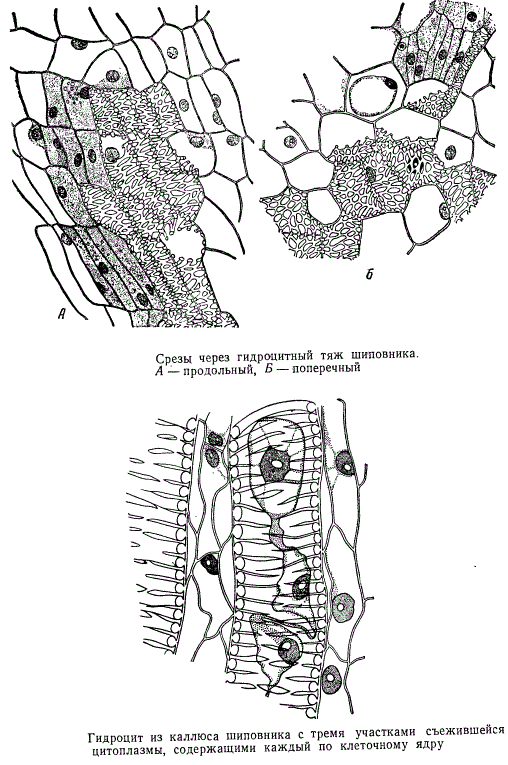
Среди гидроцитов каллюса, представляющих собой отдельные клетки, соединяющиеся друг с другом окаймленными порами сетчатого типа, есть как паренхимные, так и прозенхимные элементы.
Гидроциты каллюсовой ткани, имея на стенках одревесневшие утолщения с сетчатыми окаймленными порами, являются типичными водоносными анатомическими элементами, способствующими более быстрой подаче воды, чем обычные паренхимные клетки каллюсовой ткани, в результате чего в каллюсе формируется полноценный проводящий пучок.
Процесс превращения каллюсовых клеток в гидроциты, начинающийся в глубине клина, распространяясь, постепенно достигает участков, близких к периферии каллюса, давая ответвления по всем направлениям. В итоге недалеко от периферии каждый тяж, состоящий из гидроцитов и примыкающих к ним ситовидных трубок, оканчивается скоплением гидроцитов, окантованных слоем клеток с густым цитоплазматическим содержимым, несколько напоминающих меристему. Такие скопления гидроцитов и меристемоподобных клеток имеют вид узлов или компактных скоплений. Гидроциты в таких узлах заметно короче, чем в тяжах, идущих из глубины клина, вместе с будущими ситовидными трубками они анастомозируют с подобными же элементами соседних узлов таких расширений, образуя в общем связную систему проводящих элементов.
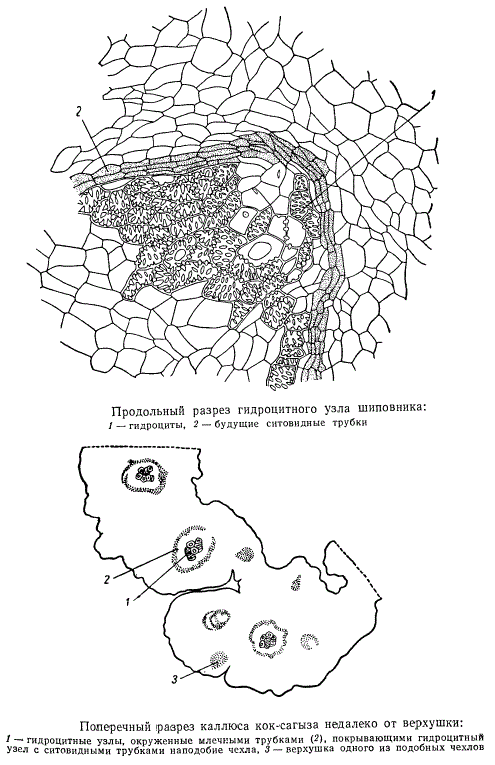
На рисунке представлен поперечный разрез через каллюс черенка кок-сагыза, проведенный несколько отступя от поверхности наплыва. В каллюсе кок-сагыза образуются такие же скопления гидроцитов (1), как и у шиповника, только у кок-сагыза они окружены кольцом будущих млечников, непосредственно примыкающих к будущим ситовидным трубкам (2). Если срез провести несколько выше узлового скопления гидроцитов, то он захватит только верхушку ткани млечников (5).
Гидроциты в каллюсе кок-сагыза образуют довольно плотную ткань, переплетаясь друг с другом в узлах. Будучи продолжительное время живыми, окруженные живыми элементами флоэмы и млечников, гидроциты представляют собой мощную систему, проводящую воду и пластические вещества, которая дает возможность быстро развиваться зачаткам побегов и корней. На «поперечном разрезе гидроцитного узла расположение составляющих его анатомических элементов не меняется по сравнению с продольными разрезами.

В мощных каллюсовых «наплывах от гидроцитных узлов первого порядка отходят тяжи гидроцитов, заканчивающиеся, в свою очередь, гидроцитными узлами второго порядка, от которых также могут отходить гидроцитные тяжи следующего порядка. В итоге в каллюсовой ткани, просуществовавшей более или менее продолжительное время, может развиться весьма сложная система гидроцитных образований.

Как было указано выше, поверхностные клетки каллюса претерпевают ряд характерных морфологических изменений, образуя ткань, подобную Пробке. При этом происходит не только отмирание и отсыхание самых наружных клеток. Часто и более глубокие слои каллюса превращаются в такую же ткань. Возможно, что отмирание наружных клеток ведет к образованию из лежащих под ними живых «леток пробковой ткани. Так как пробка принадлежит к категории защитных покровных тканей, то и ткань, образующуюся на периферии каллюса, можно назвать защитной. Защитная ткань в своем дальнейшем развитии может закладываться и в более глубоких слоях каллюса, приближаясь затем к периферии в местах выхода млечных трубок и понижаясь в промежутках между ними. Так как клетки защитной ткани образовались путем деления каллюсовых клеток и, опробковев, уже не разрастались, то они заметно мельче внутренних клеток каллюса и выделяются характерной для пробковой ткани легкой извилистостью клеточных оболочек, а также их более темной окраской.
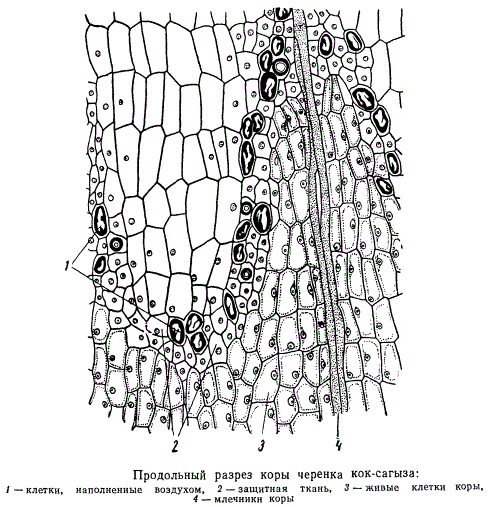
Наряду с клетками, имеющими клеточные ядра, многие клетки защитной ткани содержат только воздух. Самые клеточные ядра сильно изменены: мельче размерами, стекловидные, без ядрышек, что значительно отличает их от клеточных ядер прочих каллюсовых клеток, расположенных ковнутри от защитной ткани. В клетках, расположенных кнаружи от защитной ткани, ядра разрушаются, причем сами клетки отмирают.
Вслед за описанными структурными процессами в каллюсах начинается дифференциация почек побегов и корней из меристематических участков, структурно связанных со всеми элементами, слагающими узлы гидроцитов, и примыкающих к ним слоев. Результаты такого образовательного процесса удобнее всего наблюдать на каллюсах, культивируемых изолированно от произведших их черенков; из таких каллюсов удается выращивать молодые растеньица. Разрез каллюса, культивируемого изолированно от черенка, приведен на рисунке.
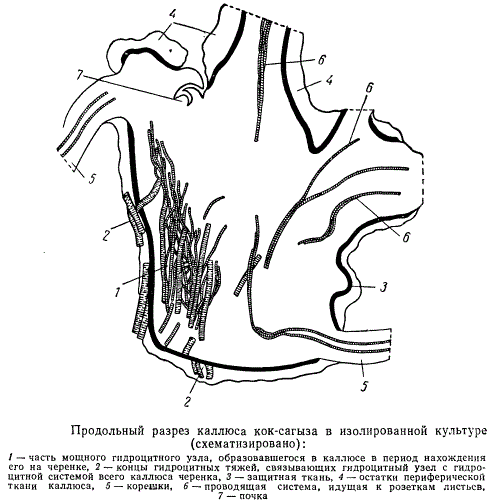
На демонстрируемом полусхематическом рисунке показан участок мощного гидроцитного узла (1), соединяющегося гидроцитными тяжами с системой гидроцитных узлов и тяжей материнского черенка. Остаток такой связи показан на рисунке цифрой 2. От гидроцитного узла отходят ветви к формирующимся почкам (7), молодым розеткам листьев (6) и корням (5). Все это покрывает паренхима и слой мощной защитной ткани каллюса (5). Защитная ткань прерывается в местах образования почек, розеток листьев и корней.
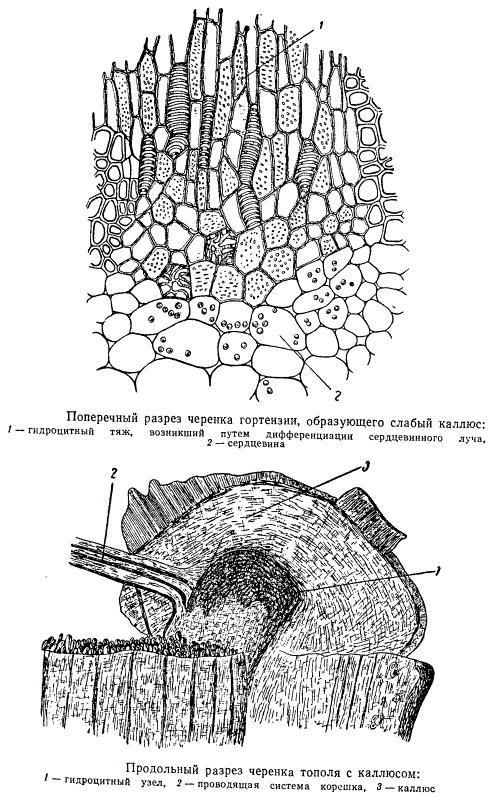
Каллюс может возникнуть из ряда тканей черенка (камбий, перицикл, сердцевина и др.), причем мощность каллюса неодинакова у разных растений. Довольно слабый каллюс образуют, например, черенки гортензии. Корни закладываются здесь, как обычно, в перицикле. От иерицикла по направлению к сердцевине клетки сердцевинных лучей дифференцируются как гидроцитная ткань, образуя перемычки с граничащими с сердцевинным лучом сосудистыми пучками.
Такой же слабый, почти незаметный невооруженному глазу каллюс образуется на черенках гардении. Корни в этих каллюсах возникают с большим трудом и таким же порядком, как у гортензии.
При развитии мощного каллюса заложение корней происходит несколько иначе. На рисунке хорошо видно место отхождения молодых корней из базального конца черенка тополя. Проводящая система корней (2) непосредственно примыкает к гидроцитному узлу, являющемуся расширением гидроцитного тяжа, идущего из каллюсового клина, образовавшегося в камбиальной зоне черенка. Почки побегов возникают на верхнем полюсе черенка. К почкам подходит проводящая система — непосредственное продолжение гидроцитного тяжа, получившего начало из клина каллюса черенка.
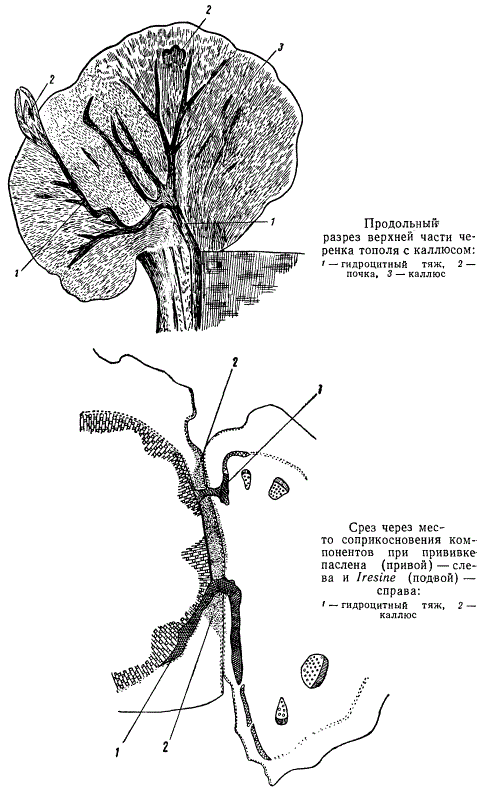
Все вышеизложенное показывает, насколько сложны структурные процессы, развертывающиеся при укоренении черенков. Еще более сложны они в подвое и привое при различных приемах прививок. Остановимся на кратком разборе явлений срастания подвоя с привоем при прививках между представителями двух различных семейств: паслена (семейство пасленовые) и ирезине (семейство амарантовые). Прививка паслена на ирезине происходит с трудом, участки срастания тканей немногочисленны, вследствие чего картина срастания сравнительно проста и удобна для изучения. Принципы структурного причленения привоя к подвою в основном сходны при всех прививках.
Наиболее существенным для удачной прививки является процесс установления контакта сосудистой системы привоя и подвоя. Подвоем в рассматриваемом примере служит ирезине, привоем — паслен. Прививка производилась сближением (копуляцией) или в расщеп.
При соприкосновении поверхностей срезов подвоя и привоя через некоторое время и при соответствующих условиях в местах срезов образуется каллюс, разрезы которого изображены на рисунках. Направо расположен подвой (ирезине), налево — привой (паслен). Обычным порядком в каллюсе дифференцируются гидроцитные тяжи (1), пронизывающие толщу каллюса (на рисунке заштрихованы точками), идущие навстречу друг другу, от привоя к подвою. Гидроцитные тяжи состоят из таких же элементов, какие описаны в гидроцитных тяжах каллюсов черенков, т. е. из трахеидоподобных клеток, коротких и длинных, с примыкающими к ним элементами флоэмы. Как при возникновении каллюса в черенках, так и при прививках, каллюс на своей поверхности покрывается защитной тканью. Последняя в нашем примере прорывается только местами, поблизости от ненарушенных чрезмерно срезом участков центрального цилиндра привоя или подвоя. В эти-то прорывы защитной ткани и направляются гидроцитные тяжи, посредством которых осуществляется связь сосудистых систем привоя и подвоя.
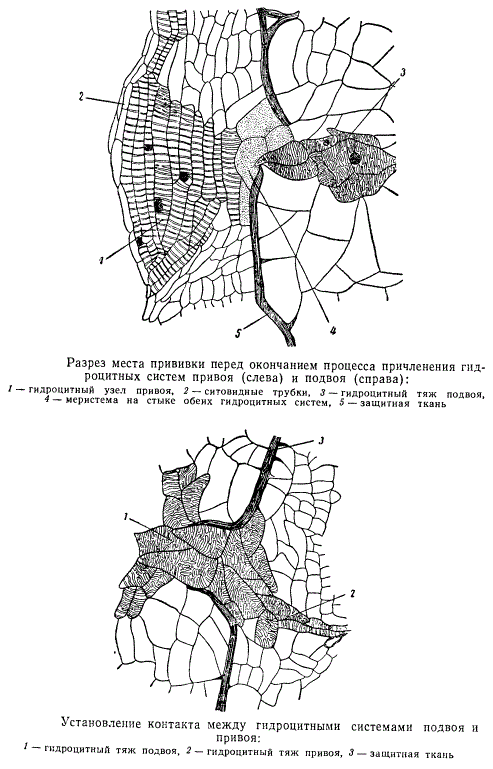
На рисунке изображена одна из таких перемычек. К мощному гидроцитному узлу привоя (1), состоящему из вытянутых элементов, с примыкающим к этим элементам слева тяжем ситовидных трубок (2), направляется гидроцитный тяж подвоя (3). На границе стыка этих двух гидроцитных систем расположены меристематические клетки (4), возникшие из каллюса привоя. В итоге при благоприятных условиях осуществляется полный контакт проводящих систем привоя и подвоя. На рисунке видно, что защитная ткань в месте прорыва отогнута в сторону подвоя; это указывает на то, что из двух идущих навстречу друг другу тяжей, дифференцирующихся в проводящие элементы клеток каллюсовой ткани, один, принадлежащий каллюсу привоя, более мощный, и дифференциация клеток в нем происходит энергичнее. Клетки каллюса подвоя заметно крупнее клеток каллюса привоя, поэтому довольно легко разобраться в местах причленения друг к другу гидроцитных систем каждого члена прививки.
Кроме причленения проводящей системы при помощи дифференцирующихся из каллюса гидроцитных тяжей — универсальных анатомических элементов высших сосудистых растений — привой и подвой приходят в соприкосновение друг с другом еще поверхностями защитных тканей каллюсов каждого компонента прививки. По-видимому, здесь срастания не происходит, осуществляется лишь слияние в единую защитную ткань облитерированных, отмерших клеток. Защитная ткань остается без коренных структурных изменений на все время существования привитого растения, срастание же сосудистой системы с течением времени расширяется и углубляется.