Способность приспособительного реагирования на условия не является универсальным свойством живой системы.
Отсюда в задачи эволюционной биологии входит изучение механизмов возникновения приспособлений, среди которых для растений наиболее типичны функциональные проявления (Ю. Сакс, К. Гебель, Г. Клебс, К. А. Тимирязев). Бесспорно, указанная проблема лучше разработана применительно к животным, однако она давно обсуждается и в физиологии растений, хотя вопрос g способах (модусах) филогенетического преобразования органов и функций растений еще слабо разработан.
Применительно к функциональным, явлениям выделены следующие количественные закономерности приспособительного реагирования организмов на изменение среды (Г. Ф. Гаузе, 1984): неизменность функций (сохранение важных функций при изменении второстепенных), выключение функций (ингибирование фермента при отсутствии субстрата), усиление функций, сочетание усиленных функций между собой. Все это в какой-то мере проявляется и у растений (К. В. Манойленко, 1984). Однако для них в первую очередь можно выделить такие модусы, как множественность реализации, усиление или ослабление, компенсация, стабилизация и автономизация функций, перекомпоновка функций.
Множественность реализаций функций. Это предпосылка филогенетического преобразования функциональных особенностей. Здесь, как и в случае преобразования органов в филогенезе, проявляются характерные для любой структуры мультифункциональность и способность любой функции изменяться количественно (А. Н. Северцов, 1939)1 По мнению А. М. Голдовского (1975), в пределах каждой группы веществ существует целый ряд близких по строению и функции соединений. Подобные наблюдения легли в основу сформулированного им закона множественности или многовариантности протекания реакций.
Множественность реализации функций подтверждается:
- возможностью изменения первичных и вторичных продуктов метаболизма;
- участием неродственных веществ в регуляции одного и того же явления (например, рост и развитие зависят от комплекса энергетически и физиологически активных веществ);
- индуцированием одной и той же реакции разными воздействиями (например, развитию ксерофильной структуры способствуют засуха, интенсивность освещения, некоторые химические агенты и т. д.).
Множественность реализации функций свидетельствует прежде всего о сложности процессов жизнедеятельности растений, она способствует повышению надежности онтогенеза за счет расширения нормы реагирования организма. Например, наличие разных ферментов, катализирующих одну и ту же реакцию (Fe- и Cu-протеины, флавопротеины и т. д.), заметно повышает интенсивность дыхания в благоприятных условиях (Б. А. Рубин, М. Е. Ладыгина, 1974). Точно так же в качестве приспособления для улавливания света В толще водоема возникает разнообразие пигментов у бактерий и водорослей (А. Клюйвер, К. Ван-Ниль, 1959; Е. Н. Кондратьева, 1972). Поэтому продукты метаболизма, состав пигментов и способы дыхания у бактерий и водорослей оказываются более разнообразными, чем у цветковых. Усиление специализации метаболических звеньев имеет значение для интенсификации процессов жизнедеятельности.
Блочный принцип эволюции или перекомпоновка функций. Для блочного принципа эволюции функций (А. М. Уголев, 1985) имеет значение мультифункциональность на уровне макромолекул, выступающая в виде их мультипотенциальности. Специализация молекул приводит к уменьшению их мультипотенциальности и интенсификации функций. Все это проявляется на молекулярном клеточном и тканевом уровнях.
Любая сложная функция возникает из менее простых компонентов. Так, сложные процессы живых организмов складываются из сочетания особенностей элементарных частиц, химических элементов и веществ, минералов (Д. Лима де Фариа, 1991). Например, на основе способности красящих веществ абиогенного происхождения (пигментов) на свету поглощать CO2 и выделять O2 с возникновением живых существ складываются их. фотохимические процессы, в том числе и фотосинтез. Вся специфика живых организмов есть ничто иное, как объединение в комплексы молекул с разным сочетанием без потери индивидуальности каждой из них (М. Эйген, 1973; С. Э. Шноль, 1979).
Подобная «химическая предопределенность» и способствует становлению жизни. Отбор в ходе предбиологической эволюции шел по принципу максимального повышения каталитической активности не только макромолекул, но и эффективности сочетания их в системах путем усиления пространственно-временной упорядоченности удачных сочетаний (В. А. Карасев, В. Е. Стефанов, 1991). Сочетание структур с мультифункциональными особенностями — предпосылка многих физиологических явлений.
Вероятно, первично и система синтеза белка происходила на основе простых каталитических систем (Р. Фокс, 1992). С образованием нуклеопротеинов усиливаются специфичность синтеза белков и разнообразие их функций благодаря отбору. Так, образование молочной кислоты и в бактериях, и в мышцах человека имеет одинаковый механизм, но роль молочной кислоты меняется (Дж. Мецлер, 1980).
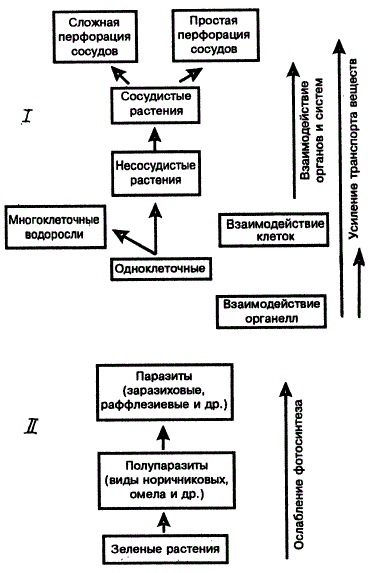
Усиление или ослабление функций имеет место при становлении механизмов фотосинтеза и дыхания. Не менее убедительные данные дает изучение эволюции процессов передвижения веществ. При переходе к многоклеточной организации усиление транспорта веществ достигается с помощью разных механизмов и развития новых структур (К. Эсау, 1969; А. Л. Курсанов, 1976).
Тенденцию усиления функций можно проследить и на примере механизмов водоотдачи. Первичноводные растения не обладали сколько-нибудь развитой системой активного испарения. Излишняя жидкость, накапливающаяся в организме, выводилась у них со всей поверхности или через специальные поры — гидатоды, как у некоторых современных наземных растений. С выходом на сушу (при высокой влажности воздуха) у растений усиливается кутикулярная, а в связи с арцдизацией климата — устьичная транспирация. Такую же смену способов транспирации проследил П. А. Генкель в онтогенезе типичного наземного растения как выражение рекапитуляции функций.
Постепенное усиление функций можно продемонстрировать и на примере эволюции способности к движениям у растений. В элементарной форме она присуща всем растениям, а в особых экологических условиях (борьба за влагу, свет, аэрацию и т. д.) эта способность была усилена отбором в одном из следующих направлений: круговые вращения стебля, лазание с помощью усиков, листьев и т. д.
Усиление главной функции часто достигается увеличением числа одноименных компонентов, участвующих в ее осуществлении. Например, интенсивность фотосинтеза зависит от количества хлоропластов и содержания в них хлорофилла, дыхание — от числа митохондрий, рост — от числа и размеров клеток и концентрации гормонов.
В процессе эволюции нередко наблюдается и ослабление функций. Выше было показано ослабление или полная утрата фотосинтетической активности у цветковых полупаразитов и паразитов. Другой пример — снижение интенсивности транспирации у суккулентов и некоторых ксерофитов, ослабление способности к движениям у цветковых даже в пределах одной и той же систематической группы, например у разных тыквенных.
Механизмы и причины ослабления функций не изучены. Не исключен отбор растений по обратным мутациям (структурным и функциональным) или выпадение какого-либо звена из сложной цепи метаболизма. Например, ослабление транспирации у суккулентов обусловлено подавлением механизма устьичных движений. Явления усиления и ослабления функций тесно связаны с процессами компенсации.
Компенсация функций. Известно много фактов, когда функция редуцированного органа не утрачивается, а выполняется другими структурами. Так, редукция листьев у части суккулентов как средство для предотвращения излишнего испарения воды могла бы иметь отрицательные последствия для выживания вида, если бы стебель не взял на себя функцию листьев — фотосинтез. Так, в пустынных районах юго-западной части США встречается Fouquieria splendens. Растение этого вида большую половину года из-за засухи остается безлистным и фотосинтезирует стебель даже в очень засушливые периоды. Поэтому его стебель имеет зеленые ткани с хлоропластами.
Н. А. Максимов (1955) показал, что у части ксерофитов интенсивная потеря воды при транспирации компенсируется ее активным поглощением из глубоких слоев почвы мощно развитой корневой системой. Недостаток воды в окружающей среде восполняется также образованием эндогенной воды в процессе дыхания (Б. Б. Вартапетян, 1970).
В ходе эволюции у растений формируется ряд компенсаторных механизмов, устраняющих нарушения функциональной целостности организма: метамерность дифференциации, способность к регенерации, гипертрофия структуры и функций оставшегося единичного органа и т. д.
В качестве примера компенсации функции укажем на «подталкивание» реакций в процессах фотосинтеза. В отличие от животных в клетках растений, в частности в хлоропластах, триозофосфатдегидрогеназа (ТДГ) связана с НАДФ ∙ Н (акцептором электронов и протонов). Активность НАДФ ∙ Н — зависимой ТДГ — определяется наличием света и приводит к образованию НАДФ ∙ Н. Выработка большего количества НАДФ ∙ Н с участием ТДГ благоприятствует «подталкиванию» реакции в сторону образования глицеральдегид-3-фосфата (ГАФ). Без такого «подталкивания» не удалось бы преодолеть термодинамического барьера реакции, катализируемой фосфоглицераткиназой: 1,3-дифосфоглицерат + АДФ → 3-фосфоглицерат + АТФ.
Из-за недостаточности свободной энергии (1 Дж/моль) может возникнуть термодинамический барьер на пути осуществления фотосинтеза. Этот барьер преодолевается накоплением большого количества АТФ (благодаря фотофосфорилированию) и расходованием 1,3-ДФГ (при участии активной НАДФ ∙ Н — зависимой ТДГ).
Эти процессы осуществляются только при компенсации функции фосфоглицераткиназы активной НАДФ ∙ Н — зависимой ТДГ.
Явление компенсации функций видно на примере изменения активности ферментов в зависимости от условий. Так, на РБФК показано, что в темноте, даже при наличии CO2 в среде, она мало активна, тогда как после включения света резко возрастает в считанные секунды. Постепенное уменьшение интенсивности света приводит к соответствующему падению активности РБФК за 2—10 мин (Ю. А. Вийль, Т. П. Пярник, 1992). Этому способствует также полное прекращение регенерации РБФ в тканях растений в темноте.
Как известно, в РЦФ использование поглощенной энергии для переноса электронов от донора к акцептору происходит нередко против термодинамического градиента. Благодаря этому сохраняется сопряжение потока энергии и потока электронов (А. Т. Мокроносов, В. Ф. Гавриленко, 1992). В целом эффективность работы РЦФ достигается непрерывной регенерацией компонентов цепи, что сопровождается их новообразованием и восстановлением электронной структуры ранее, окисленного пигмента.
Известно, что у злаков хлорофилл содержится в остьях и колосковых чешуйках. Эти структуры, находясь ближе к зерновке и в хорошо освещенном положении, играют важную роль в наливе зерна. Точно так же хлорофилл околоплодника многих растений участвует в фиксации CO2, выделяющегося тканями плода при дыхании. Показано, что хлоропласт мезокарпа плода гороха фиксирует CO2 воздуха, а внутреннего эпидермиса — до 50—70% CO2, выделяемого семенем (Дж. Эдвардс, Д. Уокер, 1986).
Присутствие хлорофилла в плодах и зародышах имеет большое значение для обеспечения их тканей продуктами фотосинтеза и эндогенным кислородом (М. С. Яковлев, Г. Я. Жукова, 1993). Например, количество хлорофилла в плодах томатов составляет всего 0,25% от содержания его в листьях; тем не менее фотосинтез плодов оказывается в пределах 4—5% от фотосинтеза листьев (Б. Б. Вартапетян, 1970). Подобные возможности расширяют норму реакции организма, повышают надежность морфогенеза и способствуют поддержанию гомеостаза системы в целом.
Стабилизация и автономизация функций. Пластичность растений хорошо известна. В то же время процессы морфогенеза и жизнедеятельности растений нельзя рассматривать как нестабильные, легко изменяющиеся. Это подтверждается данными по изучению эндогенных ритмов, полярности развития, покоя, фотопериодизма у растений и т. д.
Эволюция ряда физиолого-биохимических процессов, как и онтогенеза в целом, шла в направлении повышения их относительной независимости от внешних условий (L. Whyte, 1965). Поэтому организму присуща стабильность жизнедеятельности. Так, накопление эфирных масел, имеющих значение для реализации воспроизводительных потенций растений, мало зависит от факторов среды (А. А. Прокофьев, 1955). Многообразие метаболических путей как показатель дифференцировки на молекулярном уровне — предпосылка для стабилизации жизнедеятельности организма в постоянно меняющихся условиях среды. По этой причине лабильность отдельно взятой реакции в сложной цепи биохимических процессов не приводит к преобразованию или заметному нарушению метаболизма благодаря барьерным механизмам. Так, при недостатке CO2 в воздухе растение использует для фотосинтеза CO2, выделяемый при дыхании; при избытке CO2 в тканях ночью осуществляется карбоксилирование.
Процессы жизнедеятельности протекают по универсальному принципу обратной регуляции, т. е. накопление промежуточных продуктов служит сигналом для прекращения их синтеза, тогда как их нехватка стимулирует механизмы новообразования. Это показано при изучении фотосинтеза с использованием 2-диокси-D-глюкозы. Биосинтез хлорофилла характеризуется динамической устойчивостью в меняющихся условиях среды, так как осуществляется по принципу авторегуляции (В. Л. Калер, 1976). Эти примеры свидетельствуют об автономности физиологических процессов.
Биосинтез хлорофилла, находящийся под контролем полиферментного комплекса, регулируется сложной системой механизмов, обеспечивающих строгую координацию последовательных реакций. В нормальных условиях порфирины не накапливаются в клетке из-за соответствия между скоростью их синтеза и превращения. В этом отношении интересен пример с протохлорофиллидом, синтез которого даже в темноте продолжается лишь до определенного уровня, после чего его накопление идет на убыль ввиду прекращения синтеза предшественника. Биологический смысл такой регуляции очевиден, так как накопление порфиринов при отсутствии их превращений было бы губительным для клетки из-за высокой их фотосенсибилизирующей активности и бесполезной затраты исходного материала (А. Т. Мокроносов, В. Ф. Гавриленко, 1992). По этой причине синтез порфиринов находится под метаболическим, генетическим и фитохромным контролем, что стабилизирует механизм синтеза хлорофилла и фотоморфогенез растений.
Благодаря сложной регуляторной системе внутриклеточных метаболических процессов, а также морфогенетических взаимодействий на уровне целого организма достигается автономизация онтогенеза, закономерно усиливающаяся в ходе эволюции растений. При автономизации онтогенеза внешние условия играют роль сигналов, запускающих внутренние механизмы морфогенеза. Внешняя среда поддерживает морфогенез энергетически, но при сильных изменениях факторов внешней среды у растений возникают тераты (уродства). Однако необходимо подчеркнуть, что тераты не наследуются, что опять-таки показывает важность усиления автономизации онтогенеза в ходе эволюции.