Основные понятия
Пластиды являются органоидами протопласта, характерными только для растительных клеток. Их нет лишь у бактерий, синезеленых водорослей и, возможно, грибов.
У высших растений пластиды находятся во взрослых вегетативных клетках всех органов — в стебле, листе, корне и цветке. Пластиды — это сравнительно крупные органоиды, значительно крупнее митохондрий, а иногда даже и крупнее ядра, более плотные, чем окружающая их цитоплазма, хорошо видимые в световой микроскоп. Они имеют характерное строение и выполняют различные функции, связанные главным образом с синтезом органических веществ.
Во взрослой растительной клетке в зависимости от окраски, формы и функции различают три основных типа пластид: хлоропласта (пластиды зеленого цвета), хромопласты (пластиды желтого и оранжевого цвета) и лейкопласты (бесцветные пластиды). Последние по своему размеру меньше пластид двух предыдущих типов.
Хлоропласты
Это наиболее изученные и имеющие наибольшее значение пластиды. Они содержат зеленый пигмент хлорофилл. Этот пигмент находится в растениях в нескольких формах. Благодаря хлоропластам, точнее благодаря содержащемуся в них хлорофиллу лик земли выглядит зеленым. Среди высших растений хлорофилла нет лишь у некоторых сапрофитов и паразитов, а также у растений при содержании их в полной темноте. Такие растения имеют обычно бледно-желтую окраску. Синтез хлорофилла происходит обычно только на свету, поэтому при содержании растений в темноте они остаются незелеными и называются этиолированными. В хлоропластах содержатся также и другие пигменты, относящиеся к группе каротиноидов, в частности желтый — ксантофилл и оранжевый — каротин, но обычно они маскируются хлорофиллом. По химическому составу хлоропласта несколько отличаются от цитоплазмы: например, содержание липидов в хлоропластах составляет 20—40% от сухого веса, тогда как в цитоплазме их всего 2—3%.
Структурной основой хлоропласта являются белки (около 50% сухого веса), они содержат также 5—10% хлорофилла и 1—2% каротиноидов. Как и в митохондриях, в хлоропластах обнаружено небольшое количество РНК (0,5—3,5%) и еще меньше ДНК. Исключительное значение хлоропластов в том, что в них происходит процесс фотосинтеза. Крахмал, образующийся при фотосинтезе, называется первичным, или ассимиляционным, он откладывается в хлоропластах в виде мелких крахмальных зерен. Для нормального протекания фотосинтеза необходимо присутствие хлорофилла. Хлорофилл — главное действующее начало в осуществлении фотосинтеза. Он поглощает энергию света и направляет ее на совершение фотосинтетических реакций. Из пластид хлорофилл можно извлечь с помощью спирта, ацетона или других органических растворителей. Роль желтых пигментов в фотосинтезе еще недостаточно выяснена. Предполагают, что они также поглощают солнечную энергию и передают ее хлорофиллу или же вместе с ним осуществляют специфические, важные для фотосинтеза реакции.
В соответствии с их функциями хлоропласты находятся преимущественно в фотосинтезирующих органах и тканях, обращенных к свету — в листьях и молодых стеблях, незрелых плодах. Иногда хлоропласты встречаются даже в корнях, например, в придаточных корнях кукурузы. Но основное их количество сосредоточено в клетках мезофилла (мякоти) листа.
В отличие от других органоидов, хлоропласты высших растений характеризуются однообразием и постоянством формы и размеров. Чаще всего они обладают дискообразной или линзовидной формой и когда лежат плашмя, имеют округлые или многоугольные очертания. В этом случае их часто называют также хлорофилловыми зернами. Размер хлоропластов довольно постоянен и даже у разных видов высших растений колеблется в незначительных пределах, составляя в среднем 3—7 мк (толщина 1—3 мк). Более крупные хлоропласты у высших растений встречаются редко. Например, у селагинелл (плауновидных) в клетках кожицы листьев встречаются один-два крупных хлоропласта пластинчатой формы. Величина и форма хлоропластов изменяются в зависимости от внешних условий. У растений тенелюбивых хлоропласты в общем крупнее, чем у светолюбивых, и, как правило, более богаты хлорофиллом. Обычно клетка несет большое количество хлоропластов, и число их сильно меняется; в среднем же в ней насчитывается от 20 до 50 хлоропластов. Особенно богаты хлоропластами листья, а также молодые незрелые плоды. Общее количество хлоропластов в растении может быть громадным; например, во взрослом дереве насчитываются десятки и сотни миллиардов хлоропластов. Число хлоропластов в клетке связано с их величиной. Так, у кукурузы в клетках листьев обычно содержится по нескольку хлоропластов, но у сортов с особенно крупными хлоропластами число их в клетке снижается до двух.
У многих низших растений (водорослей) форма, число и размеры хлоропластов весьма разнообразны. Они могут иметь пластинчатую форму (Mougeotia), звездчатую (Zygnema) или быть в виде спиральных лент (Spirogyra) и ребристых цилиндров (Closterium). Такие хлоропласты обычно очень крупны, встречаются в клетке в небольшом количестве (от одного до нескольких) и называются хроматофорами. Но и у водорослей могут встречаться хлоропласты обычной линзовидной формы, и в этом случае число их в клетке обычно велико.
В клетках высших растений хлоропласты расположены в цитоплазме таким образом, что одна из их плоских сторон обращена к оболочке клетки, причем особенно много их около межклетников, заполненных воздухом. Здесь они тесно прилегают друг к другу и очертания их становятся угловатыми. Однако положение хлоропластов в клетке может меняться в зависимости от внешних условий и прежде всего освещенности. Они располагаются в клетке так, что улавливают свет наилучшим образом, не подвергаясь вместе с тем действию прямых солнечных лучей. В листьях некоторых растений на рассеянном свету хлоропласты располагаются преимущественно на тех стенках клеточной оболочки, которые обращены к поверхности органа, на ярком же свету они сосредоточиваются на боковых стенках или поворачиваются к лучам узкой стороной, т. е. ребром. Такое же передвижение хлоропластов наблюдается иногда и под влиянием других раздражителей — температурных, химических, механических и др. Является ли перемещение пластид активным или пассивным (током цитоплазмы), до конца еще не выяснено, но в настоящее время больше аргументов в пользу активного перемещения.
Исходя из сложности процессов фотосинтеза, состоящего из целого ряда реакций, каждая из которых катализируется особым ферментом, можно предположить, что хлоропласты имеют упорядоченную и сложную структуру. И действительно, уже в обычном световом микроскопе часто видно, что хлоропласты не являются совершенно гомогенными, а в них наблюдаются более темные мелкие зернышки, ориентированные параллельно поверхности хлоропласта, которые были названы гранами. Исследования с помощью электронного микроскопа подтвердили существование гран и показали, что весь хлоропласт в целом и граны имеют сложную структуру.
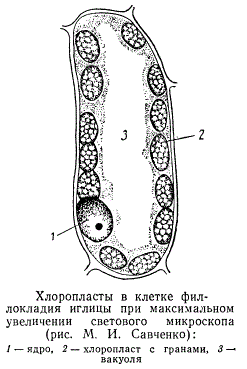
Хлоропласты в клетке филокладия иглицы
Как и митохондрии, хлоропласты представляют собой мембранные структуры, свободно лежащие в цитоплазме. От цитоплазмы они отграничены двухмембранной оболочкой с ясно видимым светлым промежутком между мембранами. Мембраны эти, как предполагают, являются гладкими и не содержат прикрепленных частиц. До недавнего времени считали, что оболочка хлоропластов сплошная, не имеет отверстий и что ее мембраны не соединены с мембранами эндоплазматической сети. Но сейчас накапливаются данные, показывающие, что это не всегда так. Иногда в отдельных местах оболочки могут возникать субмикроскопические отверстия. Возможно, что в отдельные периоды деятельности хлоропластов эти «поры» состоят в тесном контакте с эндоплазматической сетью, однако этот контакт кратковременен. Оболочка хлоропласта, обладая свойством избирательной проницаемости, играет регулирующую роль в обмене веществ между цитоплазмой и хлоропластом.
Тело хлоропласта пронизано системой двухмембранных пластин, называемых ламеллами. Пространство между ламеллами заполнено водянистой белковой жидкостью — стромой, или матриксом хлоропласта. В строме могут находиться крахмальные зерна, капли масла и рибосомоподобные частицы. Совсем недавно с помощью особо тонких методов приготовления препаратов в строме хлоропластов некоторых растений были обнаружены скопления параллельных фибрилл диаметром 80—100 Å и длиной свыше 1000 Å. Эти пучки микрофибрилл хлоропластов получили название стромацентров. Их функция совершенно не выяснена.
В отдельных участках хлоропласта ламеллы довольно плотно подходят друг к другу, располагаясь параллельно его поверхности, в результате чего в этих участках образуются скопления ламелл, называемые гранами. Внутри граны парные мембраны ламелл сливаются по краям, образуя замкнутые уплощенные мешочки, называемые дисками, или тилакоидами. Пачки таких дисков и образуют грану. Отдельные граны связаны между собой в единую систему с помощью ламелл, пронизывающих межгранные пространства. Хлорофилл не рассеян диффузно по хлоропласту, а сосредоточен в ламеллах, как предполагают, в виде мономолекулярного слоя. Рибосомы находятся не только в матриксе, но могут встречаться и на поверхности гран.
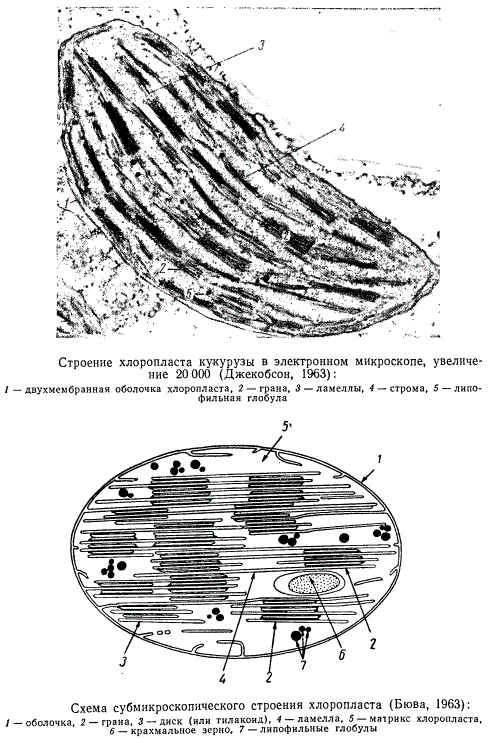
Схема хлоропласта кукурузы и схема строения хлоропласта
Число дисков в гранах колеблется от двух до нескольких десятков, а диаметр в зависимости от вида растения — от 0,3 до 2 мк. Поэтому у многих растений граны не видны в световой микроскоп. Число и расположение гран в хлоропласте зависят от вида растения, возраста и активности хлоропластов. В хлоропластах Aspidistra гран так много, что они соприкасаются между собой, а у так называемых клеток-спутников тыквы основной объем хлоропласта занят стромой. В хлоропластах листьев томата и хризантемы граны беспорядочно разбросаны, а в хлоропластах табака они правильно ориентированы по отношению к поверхности хлоропласта и располагаются на равном расстоянии друг от друга. У светолюбивых растений граны мельче, чем у теневыносливых.
Структура хлоропластов высших растений прекрасно приспособлена к выполнению их главной функции — фотосинтеза. Уже само разделение хлорофиллоносного аппарата на мелкие пластиды означает громадное увеличение активной поверхности. За счет образования мембран и гран эта поверхность увеличивается еще более. Большая активная поверхность и тонкая пространственная ориентация обеспечивают легкий доступ энергии кванта света и возможность переноса этой энергии к химическим системам, участвующим в фотосинтезе. Принцип замкнутых камер — тилакоидов, благодаря пространственному разобщению позволяет одновременно и независимо осуществлять один и тот же комплекс реакций, составляющих фотосинтез. В рибосомах хлоропластов идет синтез белка.
В клетках некоторых водорослей (спирогира) и реже высших растений (например, клетки так называемой обкладки проводящих пучков у кукурузы) встречаются безгранные хлоропласты, у которых ламеллы пронизывают строму, не образуя отчетливых гран.
Происхождение и развитие хлоропластов изучено еще очень мало и единой точки зрения по этому вопросу пока не существует. Известно, что в молодых, эмбриональных клетках дифференцированных хлоропластов нет. Вместо них имеются так называемые пропластиды. Это очень мелкие (доли микрона) тельца, находящиеся на грани разрешающей способности светового микроскопа. Первоначально они имеют амебовидную форму (несут лопасти), отграничены от цитоплазмы двойной мембраной и не содержат ни внутренних мембран, ни хлорофилла. Внутренние мембраны, образующие ламеллы, развиваются позднее. Имеется несколько гипотез о дальнейшем развитии пропластид. Согласно одной из них, в прозрачной строме пропластиды сначала образуются скопления мельчайших пузырьков, расположенных в правильном порядке наподобие кристаллической решетки. Это скопление пузырьков, каждый из которых одевается собственной мембраной, называется первичной граной. По периферии первичной граны возникают ламеллы, которые распространяются во все стороны от нее. В дальнейшем на них образуются все ламеллярные структуры хлоропласта, в том числе и граны. На свету в них наблюдается отложение пигментов и, прежде всего, хлорофилла.
По другой гипотезе, ламеллы первоначально образуются как складки внутренней мембраны оболочки пропластиды, а не из пузырьков. При этом сначала возникает структура, подобная митохондрии.
Таким образом, эти гипотезы исходят из принципа непрерывности пластид и отрицают их происхождение из других органоидов протопласта и, прежде всего, из цитоплазмы. Однако другие ученые считают, что митохондрии и пластиды по происхождению тесно связаны между собой. Например, удалось показать возникновение митохондрий из зрелых хлоропластов путем «почкования». Впоследствии эти митохондрии могли снова объединяться с хлоропластами. Но все эти гипотезы не получили еще достаточного обоснования, и вопрос о происхождении пластид еще ждет своего решения.
Кроме возникновения из пропластид, хлоропласты могут размножаться путем простого деления. При этом из взрослого хлоропласта образуются две дочерние пластиды, часто неравных размеров. Электронномикроскопическая картина такого деления до сих пор не изучена.
Структура хлоропласта не остается постоянной, она закономерно изменяется в процессе роста клетки. Изменение структуры хлоропластов с возрастом листьев заметно даже в световой микроскоп. Так, молодым листьям обычно соответствует тонкогранулярная структура, листьям среднего возраста — крупногранулярная структура. В стареющих листьях происходит нарушение структуры и деградация хлоропластов.
Хлоропласты — довольно нежные непрочные органоиды. При помещении клетки в дистиллированную воду или гипотонический солевой раствор они быстро разбухают, на их поверхности образуются пузыревидные вздутия, а затем они расплываются. Электронномикроскопические исследования показали, что разбухание происходит в строме, а не в ламеллах. При повреждении клетки хлоропласты, рассматриваемые в обычный микроскоп, сначала становятся грубогранулярными, вздуваются, приобретают пенистый вид и, наконец, гранулярность исчезает. Патологические изменения происходят в хлоропластах листьев и при недостатке в почве элементов минерального питания. Однако хлоропласты некоторых клеток могут обнаруживать и высокую стойкость. Так, у деревьев зеленый цвет коры обусловлен наличием слоя клеток с хлоропластами. Эти хлоропласты прекрасно переносят низкие температуры и переходят в активное состояние, обнаруживаемое по сильному позеленению коры, например, у осины, очень рано весной, когда ночью еще бывают сильные морозы. Низкие зимние температуры переносят также хлоропласты листьев (хвои) наших вечнозеленых хвойных деревьев. При этом, как показали электронномикроскопические исследования, они сохраняют свою сложную внутреннюю организацию.
Лейкопласты
Это мелкие бесцветные пластиды. В световом микроскопе их часто трудно обнаружить, так как они бесцветны и обладают тем же коэффициентом преломления, что и цитоплазма. Обнаружить их можно лишь в случае накопления внутри них крупных включений. Это очень нежные органоиды и при приготовлении срезов живого материала разрушаются даже более легко, чем хлоропласты. Они встречаются во взрослых клетках, скрытых от действия солнечного света: в корнях, корневищах, клубнях (картофель), семенах, сердцевине стеблей, а также в клетках, подвергающихся сильному освещению (клетки кожицы). Часто лейкопласты собираются вокруг ядра, окружая его иногда со всех сторон. Форма лейкопластов очень непостоянна, чаще всего это шаровидные, яйцевидные или веретеновидные образования.
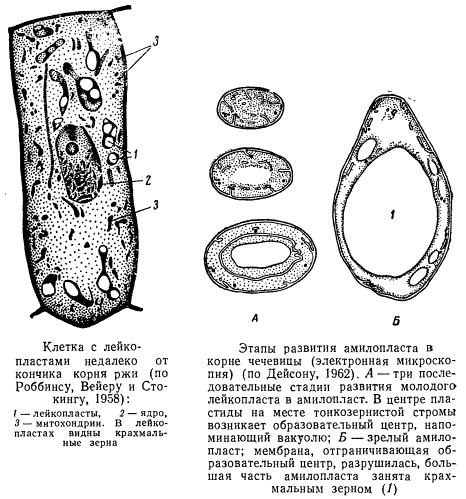
Клетка с лейкопластами и этапы развития амилопласта
Лейкопласты — органоиды, связанные с образованием запасных питательных веществ — крахмала, белков и жиров. Деятельность лейкопластов специализирована: одни из них накапливают преимущественно крахмал (амилопласты), другие — белки (протеопласты, называемые также алейронопластами), третьи — масла (олеопласты). Лейкопласты клеток кожицы листьев и стеблей нельзя отнести ни к одному из этих типов, так как функция их еще не выяснена.
Амилопласты накапливают крахмал в виде так называемых крахмальных зерен. Это преобладающий тип лейкопластов. Структура амилопластов и механизм образования крахмала с трудом поддаются изучению в световом микроскопе, а в электронном микроскопе исследованы еще слабо. Как предполагают, они образуются из пропластид, но в отличие от хлоропластов развитие их структуры не идет далеко, а задерживается на довольно ранней стадии — стадии незрелой, слаболамеллярной пластиды. Снаружи амилопласты отграничены двухмембранной оболочкой. Внутри пластида заполнена тонкогранулярной стромой. Образованию крахмальных зерен в амилопластах предшествует развитие мельчайших пузырьков, которые сливаются, уплощаются, ограничивая мембраной участок стромы в центре пластиды. Этот участок, получивший название образовательного центра, становится более светлым, напоминая вакуолю. В образовательном центре и начинается отложение крахмала. Когда будущее крахмальное зерно начинает увеличиваться в размерах, мембраны, отграничивающие образовательный центр, исчезают, в дальнейшем рост зерна идет вне связи с ними. При отложении крахмала оболочка амилопласта и строма могут сильно растягиваться, в результате чего размер амилопласта сильно увеличивается за счет растущего крахмального зерна. Крахмальное зерно заполняет затем почти всю полость амилопласта, оттесняя его живое содержимое на периферию в виде тончайшей пленки на поверхности зерна. Во многих случаях крахмальное зерно может достигать таких размеров, что амилопласт разрывается и сохраняется только на одной стороне крахмального зерна. В этом случае новые порции крахмала могут синтезироваться лишь в тех участках, где крахмальное зерно остается в контакте с мембранами и стромой амилопласта.
Развиваясь из пропластид, амилопласты при определенных условиях могут превращаться в пластиды других типов. Если, например, поместить корень ячменя на свет, то можно видеть, что некоторые лейкопласты увеличиваются в объеме и превращаются в хлоропласты, сходные с теми, которые образуются в листьях. Если такой корень вновь лишить света, то эти хлоропласты уменьшаются в размерах и теряют свой хлорофилл, но не превращаются опять в лейкопласты, а вырабатывают глобулы (шарики) каротина, становясь таким образом хромопластами. Олеопласты, т. е. лейкопласты, образующие главным образом масла, встречаются значительно реже, чем амилопласты (например, в клетках листьев некоторых однодольных). Они обычно представляют собой продукт старения хлоропластов, которые теряют хлорофилл. При этом в строме пластиды возникают мельчайшие глобулы масла. Затем пластидная оболочка разрушается, и содержимое соседних пластид сливается, образуя более крупные жировые капли. Иногда в таких пластидах одновременно накапливается и крахмал.
Синтез запасного белка — протеина — может осуществляться в третьем типе лейкопластов — протеопластах. Белок в виде кристаллов и зерен образуется в семенах многих растений, особенно тех, которые содержат еще и много масла (например, семена клещевины). Протеопласты, подобно амилопластам, возникают из пропластид. Их развитие также задерживается на стадии незрелой ламеллярной пластиды. В строме протеопласта запасной белок первоначально накапливается в виде фибрилл, которые затем объединяются в более крупные пучки. Далее оболочка и строма пластиды разрушаются, а пучки белковых фибрилл превращаются в подобие мелких вязких вакуолей. Затем вакуоли соседних пластид сливаются, часть белка оформляется в виде кристаллоидов.
Таким образом, и крахмал, и запасной белок, и капли масла являются инертными включениями, продуктами жизнедеятельности пластид. Причем каждый из них может накапливаться не только в лейкопластах, но также в хлоропластах и хромопластах. Но если крахмал образуется в пластидах, то запасные белки и жиры очень часто могут иметь и внепластидное происхождение, возникая, вероятно, непосредственно в цитоплазме и независимо от пластид. Структурные процессы, происходящие при этом, до сих пор изучены слабо.
Хромопласты
Хромопласты представляют собой пластиды желтого или оранжевого и даже красного цвета. Они встречаются в клетках многих лепестков (одуванчик, лютик, калужница), зрелых плодов (томаты, шиповник, рябина, тыква, арбуз, апельсин), корнеплодов (морковь, кормовая свекла). Яркий цвет этих органов обусловлен желтыми и оранжевыми пигментами — каротиноидами, сосредоточенными в хромопластах. Эти пигменты характерны и для хлоропластов, но там они маскируются хлорофиллом. Они не растворимы в воде, но растворимы в жирах.
В отличие от хлоропластов форма хромопластов очень изменчива и определяется их происхождением и состоянием в них пигментов, а также систематическим положением образующего их растения. В зависимости от состояния каротиноидов различают хромопласты трех типов:
- хромопласты, у которых каротиноиды откладываются в форме мелких, но видимых в световой микроскоп ни с чем не связанных кристаллов (хромопласты моркови);
- хромопласты, у которых каротиноиды растворены в субмикроскопических липоидных глобулах (лепестки лютика и алоэ);
- хромопласты, каротиноиды которых собраны в пучки, состоящие из субмикроскопических нитей и связанные с фибриллами белка (красный перец, томаты, мандарин).
В отличие от хлоропластов и лейкопластов хромопласты редко возникают непосредственно из пропластид, а обычно представляют собой результат дегенерации хлоропластов. Исключение составляют хромопласты моркови, которые возникают не из хлоропластов, а из лейкопластов или непосредственно из пропластид. Части корнеплода, не погруженные в почву и развивающиеся на свету, обычно зеленеют. Это происходит не в результате превращения хромопластов в хлоропласты, а вследствие образования хлоропластов из пропластид или лейкопластов. Хромопласты вообще не могут превращаться в другие типы пластид. Чаще всего хромопласты образуются при разрушении хлоропластов, когда последние вступают в необратимую фазу развития. Таково происхождение хромопластов 2-го и 3-го типов. При этом в хлоропластах увеличивается содержание жиров и каротиноидов, которые собираются в строме пластиды в виде субмикроскопических глобул, ламеллярные структуры исчезают, а хлорофилл разрушается. Глобулы пигмента растут, а объем стромы уменьшается, в результате глобулы могут заполнить большую часть пластиды. Округлая форма «материнского» хлоропласта при этом сохраняется. Подобный процесс деградации хлоропластов происходит, вероятно, и при осеннем пожелтении листьев и при созревании плодов. Хлорофилл в желтеющих листьях разрушается и перестает маскировать каротиноиды, которые резко выступают и обусловливают желтую окраску листьев.
В корнеплодах моркови хромопласты возникают из лейкопластов, вначале крахмалоносных, при этом в строме пластиды накапливаются каротиноиды, которые позже кристаллизуются. Крахмал исчезает по мере того, как растет концентрация каротина, пластидная масса уменьшается и ее становится трудно обнаружить. Выкристаллизовавшийся пигмент составляет преобладающую по объему часть хромопласта, поэтому форма хромопласта в конечном счете определяется формой кристаллизующегося пигмента и бывает обычно неправильной: зубчатой, серповидной, игольчатой или пластинчатой. Пластинки могут иметь очертания треугольника, ромба, параллелограмма и т. д.
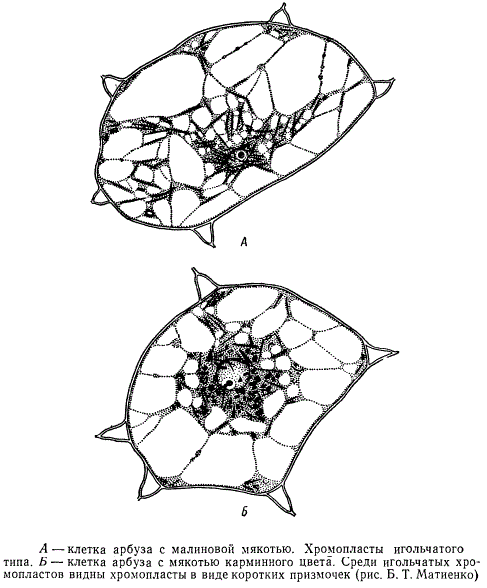
Клетка арбуза
На рисунке изображена одна из клеток арбуза с малиновой мякотью при рассматривании в световой микроскоп. В клетке видна цитоплазма, состоящая из тонких нитей, растянутых в различных направлениях. В более массивных тяжах цитоплазмы расположены игольчатые кристаллы пигмента хромопластов. Наибольшее скопление кристаллов наблюдается около ядра. У другого сорта арбуза с мякотью карминного цвета пигмент хромопластов кристаллизуется не только в виде игольчатых кристаллов, но и коротких призмочек различного размера.
Значение хромопластов в обмене веществ выяснено очень мало. Как и лейкопласты, они лишены способности к фотосинтезу, так как не содержат хлорофилла. Косвенное значение хромопластов состоит в том, что они обусловливают яркую окраску цветков и плодов, привлекающую насекомых для перекрестного опыления и других животных — для распространения плодов.