Более полное и глубокое ознакомление с процессом изменчивости тесно связано с изучением наследственности. Таким образом, наследственность будет нас интересовать в связи с проблемой изменчивости, тем более, что, как это станет ясно дальше, оба фактора диалектически взаимно связаны и составляют две, равно необходимые, стороны одного и того же процесса наследования.
Однако, наследственность будет нас интересовать и как самостоятельный фактор. Возникают вопросы о причинах способности организмов воспроизводить свойства предшествующих поколений и о закономерностях наследования. Анализ этих вопросов, в их связях с проблемой наследственной изменчивости, позволит обсудить и вопрос о значении различных форм изменчивости и наследственности в эволюционном процессе.
До работ Дарвина представления о наследственности, как определенной области знания, были фрагментарными. Какой-либо теории наследственности, строго говоря, не было. Однако, работы блестящих исследователей гибридизации (Кельрейтер, Гертнер, Шпренгель, Найт, Сажрэ, Ноден и др.) выявили некоторые закономерности наследования и были обобщены Дарвином, широко использовавшим также и свои личные наблюдения. Весь этот материал позволил Дарвину установить следующие закономерности наследования, сформулированные им еще в очерке 1844 года: «если скрещивать между собою две резко выраженные расы, потомство в первом поколении более или менее следует кому-либо из родителей или занимает совершенно промежуточное положение между ними, или же изредка принимает признаки до некоторой степени новые» (Дарвин, Соч., т. III, стр. 123). Во втором поколении, как указывает Дарвин, «потомство обычно крайне варьирует при сравнении одних особей с другими, а многие из них возвращаются почти к прародительским формам». Таким образом, еще в 1844 г. Дарвин сформулировал важнейшие закономерности наследования: 1) правило доминирования в первом поколении и 2) правило расщепления во втором поколении.
Обратимся теперь к представлениям Дарвина о причинах наследственности. Они излагаются им в его «временной гипотезе пангенезиса». В кратких чертах содержание последней таково. Дарвин принимал, что в каждой клетке любого органа содержатся мельчайшие зачатки — геммулы, которые способны к размножению и питанию, причем каждая клетка, а в конечном счете, каждый орган отделяют большое число их. Геммулы изменчивы, и их изменения возникают под влиянием внешних факторов. По Дарвину, геммулы, выделяемые отдельными клетками, разносятся по проводящим путям организма по всему телу. Обладая взаимным «сродством», геммулы собираются от всех частей тела в общий комплекс и образуют наследственную основу клеток размножения (половых в том числе). Эта гипотеза переноса геммул путем межклеточного, или интерцеллюлярного транспорта (термин, позднее предложенный де-Фризом), рассматривалась Дарвином, как объяснение явлений наследственности, в том числе — наследования вновь возникших изменений. Изменения, возникшие в органах под влиянием внешних факторов, обусловливают изменения в геммулах, а так как геммулы транспортируются в клетки размножения, то изменения геммул соответственно влияют на формообразование потомков и передаются им, т. е. становятся наследственными.
Гипотеза пангенезиса, конечно, умозрительна, спекулятивна, так как ничем не подтверждается и фактически не доказана. Отметим, однако, что в геммулах Дарвин видел не только носителей наследственности, но и те посредствующие звенья, которые обусловливают морфо-физиологическую целостность организма. Организм и его воспроизводительная система, а равно процессы изменений в них рассматриваются Дарвином в их неразрывном единстве, как целое, почему Дарвин и подчеркивал, что «в строгом смысле слова размножение не производится ни половыми элементами, ни почками, а всеми клеточками, из которых состоит организм».
Корпускулярные теории наследственности
Гипотеза пангенезиса была первой гипотезой наследственности, основанной на представлениях о морфологических единицах наследственности — геммулах. Дальнейшее развитие теории наследственности, историю которой мы кратко в дальнейшем проследим, следовало по этому же пути, т. е. базировалось на признании существования материальных носителей наследственности — корпускул наследственного вещества. Однако, методология последарвиновских теорий наследственности существенно изменилась. Уже Гальтон (1875) предложил гипотезу наследственности, в которой проводилась резкая грань между половыми и телесными зачатками. Последние обеспечивают индивидуальное развитие организма, тогда как половые клетки, по Гальтону, составляют обособленную систему, независимую от телесных клеток и факторов внешней среды. Дарвиновская идея о глубоких связях между половыми и телесными клетками и о влиянии внешних факторов на них не признавалась Гальтоном. Взгляды последнего получили развитие в последующих морфологических теориях наследственности.
Вначале 80-х годов Сакс и независимо от него Негели (1884) пришли к выводу, что не вся плазма клеток имеет значение в процессе наследования, а лишь определенная часть ее. Негели предложил различать плазму — носительницу наследственных свойств (идиоплазма) и остальную — питательную или стереоплазму. Наследственные свойства определяются, по Негели, определенным строением идиоплазмы, а именно — она состоит из мельчайших частиц, мицелл, образующих определенную структурную сеть. Каждый организм характеризуется определенной мицеллярной структурой. Всякое же изменение, возникшее в конкретном участке взаимно связанных мицелл, влечет за собою изменение целого, т. е. всей мицеллярной системы. Эта измененная система и передается потомству. В отличие от теории пангенезиса Дарвина, в теории Негели проводится идея автономности идиоплазмы, т. е. ее независимости от внешних факторов. По Негели, идиоплазма развивается в силу заложенного в ней стремления к прогрессивному совершенствованию. Точка зрения, согласно которой наследственное основание организма развивается исключительно по своим внутренним законам, вне связей с внешними факторами, должна быть названа автогенетической. Принцип автогенеза, как легко видеть, совершенно чужд дарвинизму.
Элементы автогенеза были выражены и в концепции А. Вейсмана (1834—1914), предложившей различать соматическую (сома — тело) и зародышевую плазму. Первая обеспечивает, по Вейсману, органогенез, тогда как вторая определяет передачу потомству родительских свойств.
В результате успехов цитологии и в частности выяснения процессов митотического деления клеток, а также процессов оплодотворения, была выяснена ведущая роль ядра и в особенности хромосом в процессах наследования (О. Hertwig, 1875, 1884; Strassburger, 1884; Boveri, 1889). Нашему читателю должны быть известны явления митоза, редукционного деления и оплодотворения.
Последние имеют всеобщее распространение и должны рассматриваться, как. общебиологическое явление, изучение которого способствовало укреплению и развитию идеи, что ядро имеет ведущее значение в передаче потомству наследственных свойств.
В этом направлении и формировалась теория Вейсмана (наиболее полно развитая им в 1892 г.), по которой конкретными носителями наследственности являются особые единицы зародышевой плазмы, локализованные в хромосомах. При этом Вейсман защищал идею, что видимое строение хромосом не исчерпывает их структуры. Единицами жизни Вейсман считал биофоры (носители жизни), которые слагаются в детерминанты (определители признаков). Комплексы последних образуют видимые составные элементы хромосом — иды, а иды слагаются виданты, или хромосомы.
Носителями наследственных свойств, по Вейсману, и являются детерминанты, которых в зародышевой плазме содержится столько, сколько различных структур, свойств и типов клеток содержится в организме. Следовательно, системы детерминантов и определяют наследственные особенности и признаки организмов. Проводя резкую грань между сомой и зародышевой плазмой, Вейсман установил, подобно Гальтону, тезис об автономности ее, т. е. независимости от изменений, происходящих в соме. При этом он разработал следующее общее представление об организме. Каждый новый организм претерпевает процесс онтогенеза и сам по себе приходит к концу. Наоборот, половые клетки сих зародышевой плазмой непрерывно передаются из поколения в поколение. В их рядах зародышевая плазма обеспечивает развитие новой смертной сомы и передается дальше, как таковая, последующим поколениям, никогда не образуясь заново и составляя, следовательно, непрерывный зародышевый путь.
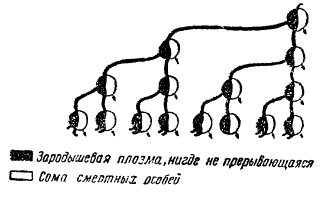
Схема зародышевого пути. Черным показана зародышевая плазма, белым цветом — сома. Зародышевая плазма непрерывна. (Из Каммерера)
По Вейсману, этот непрерывный поток зародышевой плазмы (зародышевый путь) развивается, т. е. изменяется по своим внутренним законам, определяя свойства сомы, но оставаясь независимым от нее. Что же в таком случае является источником наследственных изменений? Вейсман занимал в этом вопросе следующую позицию. Признавая теорию отбора, он распространял ее не только на формы отношений между организмом и средой, но и на отношения между частями, клетками и элементами организма.
Идея о борьбе частей была впервые высказана В. Ру (W. Roux, 1891), который защищал мысль, что между клетками, тканями и органами происходит борьба за существование.
Эту идею Вейсман и перенес на отношения в системе зародышевой плазмы. Вейсман предложил теорию герминативного отбора, т. е. отбора зародышевых элементов, детерминантов, основываясь на предположении, что детерминанты, как живые частицы, питаются за счет элементов внешней среды и растут. Так как условия питания отдельных детерминантов различны, то и темпы их роста будут не одинаковы. Энергично питающиеся детерминанты оказывают явственное влияние на соответственную часть тела. Напротив, угнетенное состояние других детерминантов сказывается и на их детерминирующем влиянии на признаки. Таким образом, от внутренней борьбы детерминантов и ее исхода и зависят, по Вейсману, индивидуальные Наследственные вариации. «Оказывается ли один детерминант в состоянии упадка или подъема, — писал Вейсман, — это во всех случаях зависит от игры внутренних сил зародышевой плазмы…» Итак, наследственные изменения возникают, по Вейсману, автономно, в силу внутренних соотношений между детерминантами.
С теорией герминативного отбора связано вейсмановское учение об амфимиксисе. Этим термином Вейсман обозначает процесс комбинирования женской и мужской зародышевой плазмы в результате оплодотворения яйца. По Вейсману, амфимиксис имеет огромное значение в формировании зародышевой плазмы потомка. Все изменения, которые произошли в системе детерминантов каждой материнской и отцовской иды под влиянием борьбы и герминативного отбора, сохраняются в зародышевой плазме потомка и влекут за собою возникновение в ней Новых комбинаций, а следовательно, и новых признаков.
Итак, по Вейсману, источниками наследственных изменений являются определяемые результатами герминативного отбора изменения в составе детерминантов, а равно их перетасовка в процессе амфимиксиса.
Методологически сходные позиции занимал также де-Фриз (de Vries, 1889), допускавший, что для каждой наследственной особенности надо принять отдельную единицу — панген. Последние, по де-Фризу, локализованы в ядре и передаются потомству по руслу зародышевого пути. Развитие организма де-Фриз связывал с предположением, что пангены проникают из ядра в плазму той же клетки, определяя ее формирование и функции, но не влияя на другие клетки. В этом положении де-Фриз видел отличие своей теории от гипотезы Дарвина. В последней допускается, что геммулы (пангены) транспортируются в половые клетки, т. е. принимается, как уже указывалось, интерцеллюлярный транспорт. В теории де-Фриза идея взаимных связей между пангенами разных клеток отрицается. Де-Фриз допускал лишь влияние пангенов на плазму той клетки, в ядре которой пангены заключены, т. е. гипотезу интерцеллюлярного (внутриклеточного) транспорта.
Изменения, возникшие под влиянием внешних факторов, не передаются половым клеткам. Последние, следовательно, обособлены, и, как было указано, изменения в них возникают автономно, в результате премутационных процессов внутреннего порядка.
Итак, в конце XIX столетия были развиты: а) идея о существовании особых морфологических единиц наследственности (геммулы, детерминанты, пангены), определяющих признаки организмов, и б) идея о независимости этих единиц от факторов внешней среды. Последняя идея знаменовала собою принципиальное отклонение от методологии дарвинизма. Дарвинизм утверждает идею глубоких связей наследственного основания с «сомой» и факторами внешней среды. В рассмотренных выше Теориях, наоборот, организм метафизически разрывается на две системы: сому и автономную зародышевую плазму.
Другой чертой описанных теорий, по крайней мере теорий Вейсмана и де-Фриза, является постановка всей проблемы наследственности в чисто морфологическом аспекте. Так как именно в ядре происходят морфологически регистрируемые закономерные явления, тогда как плазме они не свойственны, то ядро и хромосомы и стали рассматриваться как монопольный аппарат наследственности. Тот же морфологический аспект был одной из причин возникновения представлений о «наследственном веществе», как некоторой сумме отдельных морфологических частиц или «корпускул» (корпускулярные теории наследственности) — определителей признаков — и об организме, как «пучке» или мозаике признаков. Организм в таком понимании оказывается мозаикой признаков, зависимой от мозаики соответственных единиц наследственности. Таким образом, теория Вейсмана привела к чисто механистической мозаичной теории наследственности.
В дальнейшем, под несомненным влиянием Вейсмана и де-Фриза сформировалась корпускулярная теория наследственности, сущность которой и сводилась к представлению, что способность к наследственности обеспечивается передачей потомству готовых морфологических частиц, или корпускул «наследственного вещества». Проследим историю развития корпускулярной теории наследственности.
Корпускулярная теория получила поддержку на почве работ Менделя, который и рассматривается, как основоположник учения о закономерностях наследования, названного менделизмом.
Развитие корпускулярной теорий наследственности на почве менделизма
Иоганн (Грегор) Мендель (1822—1884) в 1865 г. опубликовал работу «Опыты над растительными гибридами», в которой он изложил результаты своих исследований по скрещиванию различных рас гороха (Pistim sativum).
Сформулированные им закономерности наследования вошли в науку под названием правил Менделя.
А. Правило доминирования (правило Нодена — Менделя). Если скрестить две расы гороха со взаимно противоположными признаками, то в первом поколении гибриды воспроизводят признаки одного из родительских растений. Например, при скрещивании желтосеменного гороха с зеленосеменным все формы первого поколения оказываются желтосеменными. Проявляющиеся в первом поколении признаки Мендель назвал доминантными, не проявляющиеся — рецессивными.
Проявление в первом поколении доминантных признаков и угнетение рецессивных и составляет содержание правила доминантности.
Б. Правило расщепления. Если скрестить между собою две особи первого поколения гибридов гороха, то во втором поколении происходит расщепление потомства с частичным возвратом к родительским формам. Этот процесс проявляется и в последующих поколениях.
Мендель прослеживал при этом не суммарные результаты скрещивания, а точные количественные отношения соотносительных пар признаков, образующих, по принятой терминологии, пару аллеломорфных признаков (зеленые — желтые семена, гладкое — морщинистое семя, высокий — низкорослый горох и т. п.).
Обозначим через Р родителей (Parens — родитель), через X — скрещивание, через F — поколения гибридов (Filiale generationis), через F1, F2, F3… и т. д. последовательные поколения. Пусть A, B, C и т. д. означают доминантные признаки, a, b, c и т. д. им соответствующие рецессивные. Через АА или Аа обозначается продукт скрещивания двух рас, отличимых по двум признакам одной аллеломорфной пары, т. е. зигота, полученная от копуляции двух половых клеток, или гамет. Через A и a обозначаются гаметы соответствующих рас с одним из двух аллеломорфных признаков. Из зиготы типа АА развиваются формы, наследственно однородные или гомозиготные, из зиготы Аа — наследственно смешанные гетерозиготные (термины гомо — и гетерозиготные были позднее, предложены Бэтсоном).
Опыты Менделя показали, что в F2, как сказано выше, происходит расщепление (менделирование). Если при этом прослеживается лишь одна пара аллеморфных признаков, то в среднем расщепление идет в отношении 3:1. Например, при скрещивании желтосеменной и зеленосеменной рас гороха в F1 все гибриды желтые. Следовательно, желтый горох доминирует и должен быть обозначен, как АА, тогда как рецессивный зеленый aa. В F2 расщепление идет в отношении три желтых: один зеленый (в опыте Менделя соответственно 6022 желтых и 2001 зеленых, т. е. 3,01:1). Надо было выяснить, что представляет собой это отношение. Для этой цели Мендель проанализировал состав желтых семян F2, так как было ясно, что зеленые семена этого поколения гомозиготны. Если бы это было не так, то они, в виду доминантности желтосеменного гороха, были бы не зелеными, а желтыми.
Чтобы проанализировать желтосеменной горох второго поколения, нужно было получить от него F3. Мендель высеял желтые семена F2 и вырастил из них, в условиях самоопыления, растения F3, среди которых оказались 166, давших только Желтые семена и 353 растения с желтыми и зелеными семенами. Отсюда ясно, что 166 растений с желтыми семенами были генетически чистыми доминантными (по цвету семени), а остальные 353 — гибриды. Численные отношения между ними, т. е. 353:166 = 2,13:1, а в среднем 2:1. Таким образом, анализ F2 выяснил генетический состав F3. В состав последнего, как видно из изложенного, входят: 1) генетически чистые зеленые формы (одна доля), выявившиеся еще во втором поколении и 2) желтые формы (три доли), которые в третьем поколении дали генетически чистые желтые формы (одна доля) и гибридные (две доли). Таким образом, суммарно было выяснено, что отношение 3: 1 второго поколения в действительности есть: 1 (желтые) + 2 (гибриды, внешние желтые) + 1 (зеленые), т. е. 1:2:1. Такова общая формула расщепления F2.
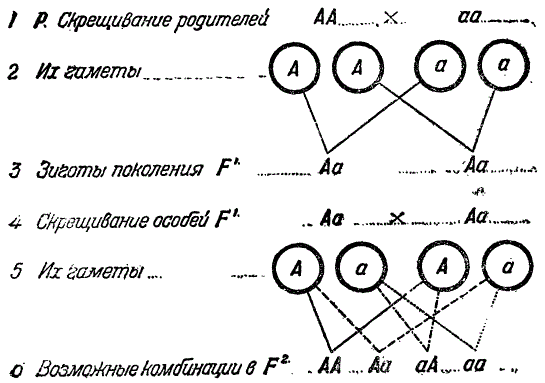
Если выразить сказанное в буквенных символах, то мы получим такую схему. Были взяты родители (Р) АА и аа. Их скрещивание дало следующие результаты.
Следовательно, состав F2 такой: AA + Aa + aA + aa(1). Отсюда видно, что при скрещивании доминантного желтого гороха с рецессивным зеленым в F1 получаются желтосеменные гибриды. Однако свойства зеленосеменной родительской фермы оказались только фенотипически подавленными, но сохраненными генотипически, и при расщеплении в F2 они снова появились (формула 1).
Из этих данных Мендель сделал вывод о вероятном существовании особых наследственных факторов, определяющих внешние признаки растения, например, желтый или зеленый цвет семян. Далее, он считал, что любые гаметы, в отношении этих факторов, остаются чистыми, например, содержат либо фактор желтого, либо зеленого цвета. Гипотеза генетической чистоты гамет основана на том, что потомство, полученное от скрещивания двух гибридов первого поколения (Аа x Аа), дает второе поколение, в котором, как мы видели, происходит расщепление с выходом форм родительского типа, т. е. АА и aa (формула 1). Стало быть, гибрид производит два типа гамет: в одних содержится только фактор А (желтого цвета), в других — только фактор a (зеленого цвета). В таком случае становится возможным свободное комбинирование факторов как результат свободной (случайной) встречи гамет разного типа, например, A♂ x A♀, или A♂ x a♀, или a♂ x A♀, наконец, a♂ x a♀, а следовательно, и расщепление потомства.
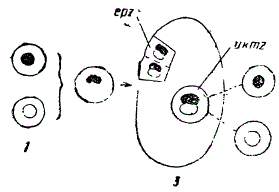
Схема, объясняющая чистоту гамет. 1 — половые клетки, отличающиеся по данному признаку („белая» гамета, „черная“ гамета). 2 — зигота, 3 — первое поколение, т. е. гибрид, epz — телесные клетки двух сортов („белые» и „черные»), ukmz — половые клетки — „белые» и „черные“ (Из Богданова)
Чтобы показать вероятность допущения существования наследственных факторов, Мендель предложил следующий контрольный опыт. Он скрестил гибрид первого поколения (Аа) с рецессивной формой (aa). Расчет здесь следующий. Если мы допустили, что гибрид Аа содержит два сорта половых клеток — A и a, то потомство, полученное от скрещивания его с рецессивной формой aa, должно дать половые клетки A и a (от гибрида Аа) и a и a (от рецессивной формы aa).
Возможные встречи между этими гаметами, очевидно, исчерпываются следующими комбинациями, которые станут ясными, если применить так называемую «решётку Пеннета»:
| A | ||
| a | Aa | aa |
| a | aA | aa |
Следовательно, должно получиться 50% гибридных желтосеменных Аа и 50% чистых зеленосеменных aa. Опыт подтвердил этот теоретический расчет, что и было истолковано, как доказательство существования в гаметах факторов зеленого и желтого цвета.
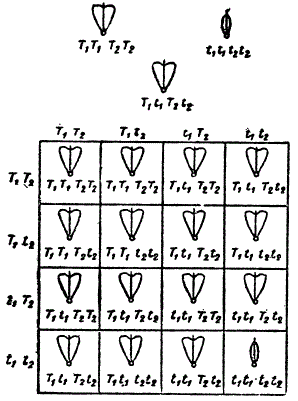
В. Правило независимого распределения зачатков. До сих пор мы имели дело с одной парой признаков, отвлекаясь от остальных, и при этом выяснили, что подобный метод возможен, несмотря на то, что скрещивающиеся формы всегда обладают многими признаками. Скрещивание, прослеженное для одной пары признаков, называется моногибридным. Возможно, однако, соответственно проследить скрещивание для двух пар признаков (дигибридное скрещивание), трех пар (тригибридное скрещивание), для многих пар (полигибридное скрещивание). Эти термины (моногибриды, дигибриды и т. д.) были предложены де-Фризом (1900).
Исследуя дигибридное скрещивание, Мендель скрещивал две расы гороха, из которых одна обладала желтыми круглыми семенами, а другая — зелеными морщинистыми. В первом поколении был получен гибрид с желтыми круглыми семенами. Следовательно, желтый цвет (А) и круглая форма (В) доминанты, a зеленый цвета и морщинистая форма b — рецессивы. Однако в F2 произошло сложное расщепление. Из 556 семян оказалось: 315 круглых желтых (А, В,) 101 морщинистое желтое (Ab), 108 круглых зеленых (а В), 32 морщинистых зеленых (ab), т. е. 9 АВ + 3 Ab + 3 aB + 1 ab. (2)
Полученные результаты показывают факт «независимого» распределения признаков. Если исходить из допущения Менделя о наследственных факторах, то придётся принять, что вопрос идет о «независимости» распределения их в половых клетках, т. е. о возможности существования четырех типов гамет, а именно: АВ, aB, Ab, ab — соответственно мужских и женских.
Это легко проверить при помощи «решётки Пеннета», в которой показаны возможные встречи между четырьмя типами мужских и женских клеток и полученные от них зиготы.

т. е. 9 желтых круглых (9 АВ), 3 желтых морщинистых (3 AB), 3 зеленых круглых (3 аВ) и 1 зеленое морщинистое (1 ab), что соответствует приведенной выше формуле (2).
Теоретический расчет, основанный на применении решетки Пеннета, подтверждается соответственными эмпирическими данными и служит иллюстрацией «независимого» распределения признаков в потомстве.
Корренс (1912) установил факт, подтвердивший наблюдения Менделя. Были скрещены две расы ночной красавицы (Mirabilis jalapa) — с белыми и красными лепестками. В F1 все цветы были розоволепестными, т. е. вместо доминирования наблюдалось промежуточное наследование. В F2 произошло расщепление обычного типа, т. е. в отношении 1:2:1, причем часть цветов была белая (1), 50% — розовые (2), остальные — красные (1). Случай этот весьма наглядно указывает на независимое распределение зачатков (факторов), хорошо выраженное фенотипически, подтверждая, вместе с тем, правило Менделя о расщеплении в F2.
Г. Цитологическое обоснование правил Менделя. Некоторое время работа Менделя оставалась почти неизвестной, пока с ней, независимо друг от друга, не ознакомились (в 1901 г.) де Фриз, Корренс и Чермак. Работая в области гибридизации и прийдя к тем же выводам, что и Мендель, они оценили значение его исследований.
С другой стороны, как уже отмечалось, в это же время были достигнуты большие успехи в области цитологии, в частности в отношении ядра и роли хромосом при делении клеток и половом процессе» Идея, что ядро является носителем наследственности, в свете работы Менделя и менделистов, приобрела более конкретные формы. При этом сложились примерно следующие представления.
Согласно механистическим воззрениям менделистов начала XX века, организм может быть представлен, как мозаика или «пучок признаков» (Гейдер, 1906). В параллель этому ядро можно трактовать как целое, слагающееся из комплекса хромосом. Далее, если, признаки образуют аллеломорфные пары, то в параллель этому хромосомы диплоидного набора образуют пары гомологичных хромосом. Если в гибридной форме комбинируются аллеломорфные признаки родительской пары, то и каждая пара гомологичных хромосом слагается из одной мужской и одной женской хромосомы. Если при расщеплении потомства в F2 появляются чистые (по данному признаку) формы, то и в половых клетках (в результате редукционного деления или мейозиса) происходит разобщение гомологичных хромосом, т. е. в каждой половой клетке остается по одной хромосоме из двух, образующих пару. Другими словами, был констатирован факт параллелизма между «поведением» хромосом и процессами расщепления. Было также показано (Сэттон, 1902, 1903), что отдельные пары гомологичных хромосом различны по форме и величине, т. е. обладают чертами индивидуальности. С другой стороны, Бовери удалось показать, что нормальное развитие яиц и осуществление всех признаков связано с нормальным набором хромосом. Следовательно, разные пары хромосом разнокачественны, подобно тому, как разнокачественны разные пары аллеломорфных признаков. Отсюда возникновение хромосомной теории наследственности, согласно которой хромосомы являются носителями наследственных свойств. Эта точка зрения натолкнулась, однако, на затруднения: хромосом всегда значительно меньше, чем признаков. Выход из затруднений был найден в предположении, что хромосомы состоят из отдельных частиц (корпускул) — факторов наследственности. Таким образом сложилась факториальная гипотеза, согласно которой отдельные единицы, или факторы, наследственности являются представителями или определителями признаков. Эти единицы наследственности получили различные наименования, но в конце концов был принят термин ген (предложенный Иогансеном), которым мы будем пользоваться в дальнейшем изложении развития учения о наследственности.
Развитие менделизма
Последователи Менделя подтвердили большое число случаев расщепления, которые в среднем соответствовали его данным.
По мере расширения опытного материала было установлено, что явления наследственности закономерны, и что эти закономерности могут быть выражены тремя основными правилами: правилом единообразия первого поколения (доминирование), правилом расцепления и правилом независимости, или самостоятельности признаков. Правила эти были догматизированы и превращены в каноны, в рамках которых рассматривалась вся проблема наследственности.
Сказанное, прежде всего, выразилось в метафизической трактовке правила единообразия (доминировании) в первом поколении. Явление доминантности, по крайней мере в течение первых двух с лишним десятилетий XX века, рассматривалось, как некоторое абсолютное свойство. Эта точка зрения особенно ярко сказалась в широком распространении так называемой теории «присутствия-отсутствия», согласно которой доминантный признак определяется соответственным, реально существующим геном (фактором), тогда как ему аллеломорфный рецессивный признак возникает вследствие отсутствия этого же фактора (гена).
Эта теория была предложена Корренсом (1902) и подробно развита Бэтсоном (1902, 1909). Она послужила способом объяснения случаев расщепления, не укладывавшихся в рамки менделевских закономерностей» Один из подобных случаев описан ниже.
У пород кур наблюдаются различные формы гребня — розовидный, гороховидный, ореховидный и простой. Первые две формы доминируют над простым, и в сочетании с ним каждая из этих двух форм дает в F2 обычное расщепление 3:1. Если же скрестить птицу с розовидным гребнем и птицу с гороховидным, то в F1 нет доминирования ни той, ни другой формы, но развиваются птицы с ореховидным гребнем. Если скрестить таких птиц первого поколения между собой, то в F2 возникает не моногибридное расщепление (как ожидалось, поскольку розовидный гребень принимался, как генетический «чистый»), а дигибридное.
При этом возникают все 4 формы гребня, указанные выше, в отношении: 9 ореховидных, 3 розовидных, 3 гороховидных, 1 простой. Как объясняется это отношение и каким образом, в частности, возник простой гребень? Было сделано предположение (Бэтсон и Пеннет, 1905), что гороховидный гребень определяется фактором Р (Pisum — горох), а розовидный — фактором R (Rosa). Фактор P превращает простой гребень в гороховидный, R — в розовидный, простой же гребень возникает вследствие отсутствия факторов R и Р. Согласно допущению, птица с гороховидным гребнем есть Pr, а птица с розовидным гребнем имеет формулу Rp где r и p означают отсутствие противоположных факторов. Отсюда возникает вывод, что ореховидный гребень появляется в первом поколении согласно следующей схеме:
Pr X Rp. .P (родители)
PR pr. . (F’).
Формы F1 могут иметь четыре типа гамет: RP, Rp, rP, rp — мужские и женские. Построив решётку Пеннета для дигибридного скрещивания, получаем:
| ♂RP | ♂Rp | ♂rP | ♂rp | |
| ♀RP | RPRP | RPRp | RPrP | RPrp |
| ♀Rp | RpRP | RpRp | RprP | Rprp |
| ♀rP | rPRP | rPRp | rPrP | rPrp |
| ♀rp | rpRP | rpRp | rprP | rprp |
т. е. 9RP + 3Rp + 3rP + rp, следовательно, 9 ореховидных, 3 розовидных, 3 гороховидных, 1 простой, возникающий вследствие отсутствия факторов R и Р.
При такой трактовке доминантность становится некоторым абсолютным свойством, а ее смена на рецессивность возникает, очевидно, не вследствие изменения свойства самого фактора (гена), а в результате его выпадения. Фактор, ген приобрел значение неизменной величины, передающейся из поколения в поколение, как константная единица наследственного вещества. Изменчивость форм превращается в игру постоянных генов, их присутствие или отсутствие.
Позднее теория «присутствия-отсутствия» пала, оказавшись несостоятельной перед лицом ряда фактов.
Одним из многочисленных доводов против теории был вскрытый исследованиями факт множественного аллеломорфизма, когда какой-либо фактор А доминантен по отношению к фактору а1, последний по отношению к фактору а2 и т. д., по общей формуле A > a1 > a2 > a3 и т. д. Было указано, что одному «присутствию» (А) не может соответствовать несколько «отсутствий» (а1, а2, а3 и т. д.). Маленькие буквы (как в разобранном примере p и r) стали символами не отсутствия, а присутствия реальных рецессивных генов. Однако и эти последние противополагались доминантным, как метафизические величины. Организм воспринимался, как слепок метафизических, неизменных зачатков. Менделизм превратился в теорию о неизменных, извечных единицах наследственности, комбинирование которых определяет богатство форм органической жизни. Эта характерная черта менделизма была по достоинству оценена Тимирязевым. Протестуя против этих «новых» идей, Тимирязев писал: «Возврат к Линнею, т. е. почти на два века назад — вот то «новое», что предлагает Бютсон взамен «старому, т. е. современному эволюционному учению…» (Тимирязев, 1916).
Дальнейшее изучение правила расщепления привело менделизм к открытию ряда случаев, которые давали в F2 необычайные расщепления, по видимости не соответствующие менделевским.
Рассмотрим здесь один из этих случаев. При скрещивании серой и белой мыши в F1 все мыши имеют серую окраску, т. е. осуществляется правило доминирования. Однако в F2 потомство этих серых мышей дает отношение: 9 серых, 3 черных, 4 белых. Чтобы свести это отношение к менделевскому 9:3:3:1, было сделано допущение, что серая окраска определяется двумя генами A и G, причем ген A (основной ген цветности) способен обусловить серую окраску лишь в присутствии гена G. Парный с последним рецессивный ген g в присутствии гена A дает черную окраску. Однако, если вместо гена A присутствует аллеломорфный ему рецессивный ген с, то мышь будет белая (aagg), так как ген a подавляет действие других генов. Сделав эти допущения, скрестим серую и белую мышь: AAgg X aagg.
Гибрид F1 от них будет (в отношении цветности) иметь формулу: AaGg и может дать следующие гаметы: AG, Ag, aG, ag (мужские и женские). Комбинация между ними дают следующий состав:
| ♂AG | ♂Ag | ♂aG | ♂ag | |
| ♀AG | AAGG серая | AAGg серая | AaGG серая | AaGg серая |
| ♀Ag | AAGg серая | AAgg черная | AaGg серая | Aagg черная |
| ♀aG | AaGG серая | AaGg черная | aaGG белая | aaGg |
| ♀ag | AaGg серая | Aagg черная | aaGg белая | aagg белая |
Как видно, четыре белых мыши представлены двумя генетическими типами: aG и ag (мыши aaGg, aagG и aagg имеют белую окраску, так как ген a подавляет проявление гена G).
Следовательно, сделанные допущения позволяют свести формулу 9:3:4 к менделевской 9:3:3:1. Было изучено большое число таких случаев с разнообразными отношениями, укладывавшимися в рамки того же приема: Из приведенного выше случая видно, что для сведения сложных случаев расщепления к менделевским допускалось реальное существование дополнительных генов, число которых подбирается в количестве, необходимом для сведения данного случая расщепления к формулам Менделя. Выработалось представление о разных типах дополнительных генов. Так, ген G в разобранном выше примере с мышами относится к группе кондициональных генов, присутствие которых необходимо, по преимуществу, для проявления той или иной окраски (пигмента), ген a относится к группе эпистатических генов, подавляющих фенотипическое проявление других генов (гипостатических). Было также разработано понятие о генах-модификаторах, воздействующих на количественное выражение других генов и т. д.
Введение понятия о дополнительных генах имело весьма важное следствие. Менделисты пришли на почве этого понятия к представлению о взаимодействии генов, образующих, следовательно, взаимодействующую систему. Эта мысль получила большое развитие и привела к следствиям, выяснение которых возможно лишь в связи с новым этапом развития генетики, известным под названием морганизма (Т. Г. Морган — основоположник современной теории гена).
Основные этапы развития морганизма
Количественно-аналитический метод Менделя и менделистов приобрел значение руководящего метода генетики. Точный гибридологический анализ по парам аллеломорфных, контрастирующих (заметно отличающихся) признаков позволил прослеживать их «судьбу» и количественные отношения. Их «независимое» распределение при расщеплении укрепило вместе с тем механистическое вейсманистское представление об организме, как мозаике признаков, зависимой от суммы наследственных единиц — факторов или генов, локализованных в хромосомах. «Мы можем принять, — писал Гейдер (1906), — что этим парам признаков, как представителям наследственного вещества, противостоят пары хромосом». Это общее положение еще не давало детализированного представления о генетической структуре хромосом и в частности о закономерностях расположения предполагаемых единиц наследственности — генов.
Факторы, или гены, трактовались абстрактно. Иогансен, предложивший самый термин «ген», не связывал его с конкретными материальными единицами.
А. Сцепление. К концу первого десятилетия XX века стали известны факты, которые открыли путь к конкретным представлениям о наследственных факторах. Некоторые признаки не подвергаются при расщеплении потомства свободному распределению, оставаясь связанными, как бы сцепленными. Явление это, получившее название сцепления, было подробно изучено Морганом и его школой и привело к созданию моргановской теории гена.
Построение этой теории связано с использованием в качестве основного объекта генетических исследований плодовых мух (Drosophila melanogaster и другие виды). Дрозофилы быстро плодятся, легко размножаются в лабораторных условиях, изменчивы в частных признаках, обладают относительно простым набором немногочисленных (8) хромосом, с ярко выраженной индивидуальностью. Исследования явлений сцепления у дрозофил показало, что число групп сцепленных признаков для каждого вида определенно и равно гаплоидному числу хромосом (n). Так, у Dr. melanogaster гаплоидный набор хромосом равен четырем хромосомам, и соответственно имеется четыре группы сцепленных признаков. У Dr. willstoni — 3 группы сцепленных признаков, а гаплоидное число хромосом n — 3. У Dr. viridis — 6 групп сцепленных признаков, n = 6, у Dr. obscura — 5 групп сцепления, n — 5 и т. д. Эти факты были объяснены тем, что каждая группа сцепленных признаков определяется генами, связанными в одной хромосоме. Поскольку хромосомы перераспределяются в гаметах и вновь комбинируются в зиготе, как некое целое, постольку заключенные в них гены, а следовательно, и представляемые ими признаки остаются связанными и не разъединяются. Следовательно, свободное (независимое) разделение признаков может иметь место только в отношении тех из них, которые зависят от генов, находящихся в разных хромосомах. Именно поэтому наблюдается соответствие числа групп сцепленных признаков числу хромосом гаплоидного набора. Эти правильные количественные соотношения между группами признаков и числом хромосом были названы законом Моргана.
Закон Моргана связывает, таким образом, определенные группы признаков, с определенными хромосомами и вместе с тем показывает, что правило Менделя о «независимом» распределении признаков не может считаться всеобщим. Это правило осуществляется только по отношению к признакам, зависимым от генов, лежащих в различных хромосомах.
Б. Кроссинговер и линейное расположение генов. Морганом было, далее, обнаружено, что признаки, обычно сцепленные, в некотором числе случаев, вновь разъединяются и подчиняются, следовательно, правилу Менделя о независимом распределении их.
Рассмотрим для пояснения следующий пример Моргана. Если скрестить самца Drosophila с двумя рецессивными признаками — желтыми крыльями и белыми глазами — с самкой, имеющей доминантно выраженные признаки, а именно — серые крылья и красные глаза, то в F1 появятся особи с серыми крыльями и красными глазами. Самка этого первого поколения была скрещена с самцом, обладавшим желтыми крыльями и белыми глазами. От этих форм было получено поколение (F2), в котором оказались: 1) мухи с желтыми крыльями и белыми глазами, 2) с серыми крыльями и красными глазами, 3) с желтыми крыльями и красными глазами, 4) с серыми крыльями и белыми глазами. В первых двух группах сохранились родительские комбинации сцепленных признаков: «желтые крылья — белые глаза», «серые крылья — красные глаза». Таких особей в F2 оказалось 99%. Остальных (3 и 4) — всего 1%, причем у этих особей сцепление указанных выше признаков нарушено, так как желтые крылья сочетались с красными глазами, а серые крылья с белыми глазами. Эти новые комбинации показывают, что сцепление между «желтыми крыльями — белыми глазами», а также «серыми крыльями — красными глазами» в 1% потомства было нарушено.
Если мы приняли, что гены сцепленных признаков находятся в одной хромосоме, то нарушение сцепления и обмен признаками может быть понято таким образом, что представляющие их гены также разъединились, т. е. переместились из одной хромосомы в другую. В нашем примере «ген серых крыльев» был сцеплен в одной хромосоме с «геном красных глаз», а «ген желтых крыльев» с «геном белых глаз». Затем в 1% поколения произошел обмен: «ген серых крыльев» сочетался в одной хромосоме с «геном белых глаз», а «ген желтых крыльев» с «геном красных глаз» — в другой хромосоме.
Возникает вопрос, как осуществляется механизм этого перемещения генов из одной хромосомы в другую? Морган сделал предположение, что гены расположены в хромосомах в линейном порядке и, что хромосомы перекрещиваются и обмениваются своими участками.
Следовательно, прежний комплекс генов разрывается, а стало быть, нарушается и сцепление определяемых ими признаков, которые вступают в новые комбинации. Это явление обмена генами, возникающее вследствие перекреста хромосом, и последующая перекомбинация ранее сцепленных признаков были названы кроссинговером (crossing over), который удалось непосредственно наблюдать. Так, воздействием рентгена Штерн получил у дрозофилы гомологичные, но разные по форме (гетероморфные) хромосомы. При перекресте этих хромосом можно было явственно наблюдать обмен их участками согласно схеме, показанной на рисунке.
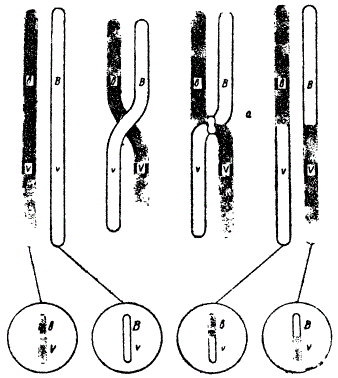
Схема кроссинговера
Морган констатировал, далее, что процент кроссинговера колеблется. В разобранном выше примере процент кроссинговера равен 1. Однако он может быть значительно выше (например, 33%, 40%).
На почве теории линейного расположения генов колебание процента кроссинговера получило следующее объяснение. Если два гена (действие которых проявляется в двух сцепленных признаках) расположены близко друг к другу один за другим, то вероятность разрыва хромосомы между этими генами меньше, чем в случаях, когда гены разделены большим расстоянием. Таким образом, процент кроссинговера зависит от расстояния между генами и, очевидно, прямо пропорционален ему. Следовательно, процент кроссинговера может служить мерилом относительных расстояний между генами, лежащими в одной хромосоме. Очень большое число исследований колебаний процента кроссинговера, сопоставленных с изменениями в самих хромосомах, привело к возможности расшифровки представлений о генном составе хромосомы и к приблизительному определению местоположения (локуса) генов в ней. Методика этого определения излагается в курсах генетики. Мы же здесь прослеживаем только развитие идей морганизма. Эти исследования затем были детализованы на почве сопоставления изменений в признаках с изменениями в строении хромосом, известными под названием хромосомных перестроек. Были выяснены следующие основные типы этих перестроек:
- транслокации, когда участок одной хромосомы отрывается и соединяется с другой хромосомой;
- инверсии, когда один участок (плечо) хромосомы перекручивается по отношению к другому ее участку (плечу) на 180°;
- делеции, или нехватки, когда средний участок хромосомы разрушается (например, под влиянием облучения), а остающиеся концы соединяются с последующим общим укорочением хромосомы;
- дупликации или удвоение какого-либо плеча хромосомы.
Во всех этих случаях было показано, что описанные перестройки хромосом в конкретных их участках связаны с изменениями определенных признаков.
Все эти факты (транслокации, инверсии, делеции, дупликации) укрепили представление, что признаки определяются линейно расположенными генами, и что по изменениям признаков и их соотношений, сопоставленным с изменениями в хромосомах, можно определять относительные местоположения или локусы конкретных генов. Отсюда возник и основной метод генетического анализа — обязательное сопоставление данных цитологического (цитогенетика) и гибридологического анализа. Огромное число исследований этого рода, предпринятых по отношению к одному и тому же биологическому объекту (Drosophila), позволило конкретизировать представление о том, 1) каков генный состав хромосом, 2) какие именно хромосомы ответственны за данные группы сцепленных признаков и 3) в каких точках каждой хромосомы расположены гены, ответственные за отдельные признаки. Кропотливые исследования этого рода позволили составить так называемые «карты хромосом», в которых показаны локусы генов, определяющих конкретные признаки.
«Парадным» (выражение Рыжкова) примером теории гена является наследование, сцепленное с полом. Было установлено, что определенные признаки наследуются в сцеплении с мужским или женским полом. Явление это было изучено параллельным гибридологическим (по признакам) и цитологическим анализом. Оказалось, что, например, хромосомный набор дрозофилы содержит два рода хромосом. Одни из них содержат гены, определяющие только общие для обоих полов признаки (аутосомы), другие, помимо таких признаков, определяют пол (гоносомы). Гоносомы дрозофилы в одних случаях одинаковы и составляют пару хромосом xx. В других случаях одна из них соответствует х-хромосоме, другая, несколько отличная по форме, обозначается как y-хромосома. Следовательно, в этом случае гоносомы образуют пару xy. Сопоставление гибридологического и цитологического анализов показало, что в случаях, когда сперматозоид содержит x-хромосому (в яйцах всегда содержатся только x-хромосомы), возникает зигота xx, которая развивается в самку. Если же сперматозоид содержит y-хромосому, то из зиготы xy развивается самец. Таким образом, стало ясным, что хромосомы связаны с процессами, определяющими пол, и что фенотипический (гибридологический) анализ совпадает с цитологическими данными. Было также установлено, что с полом сцеплены многочисленные другие признаки, следовательно, и в отношении этих последних устанавливается связь между ними и определенными хромосомами.
Особый интерес представляет явление нерасхождения гоносом, обнаруженное Бриджсом (1915). Бриджс скрещивал красноглазых дрозофил самцов с белоглазыми самками. В этих случаях в F1 возникают белоглазые самцы и красноглазые самки. Однако иногда выщепляются красноглазые самцы и белоглазые самки. На рисунке схематично показано, как Бриджс объяснял это явление. Бриджс предположил, что белоглазая самка возникает из яйца, содержавшего две x-хромосомы, которые вследствие нерасхождения остались в нем, вместо одной, как должно быть нормально. Такое яйцо имеет, следовательно, формулу xx. Оплодотворенное y-сперматозоидом, оно приобретает формулу xxy. Самка с лишней x-хромосомой и имеет белые глаза. Прямые цитологические наблюдения над этими самками показали, что они действительно имеют две x-хромосомы. Этот случай является одним из наиболее изящных доказательств наличия соотношений между хромосомами и признаками.
В. Общая концепция морганизма. Описанные выше факты и соотношения привели к следующим представлениям о механизме наследственности: 1. В хромосомах локализованы материальные единицы наследственности — гены. 2. Гены расположены в хромосомах в линейном порядке. 3. В каждой точке хромосомы при ее расщеплении (из родительского гена) образуются два дочерних гена. 4. При разделении материнской клетки происходит разъединение парных генов, которые расходятся поровну в дочерние клетки. 5. Следовательно, каждая из них получает количественно и качественно равный набор генов. 6. Любой ген происходит от гена же через удвоение материнского гена (3). Гены потомков — это гены, переданные им от родителей, через половые (и вообще пропагативные) клетки. 7. Первичный комплекс хромосом и генов, составляющих генетический аппарат зиготы, в результате ее последующего развития (например, дробления оплодотворенного яйца) и роста организма, распространяется — путем непрерывного деления клеток — на все части и органы. Поэтому в любой клетке данного организма имеется однотипный набор хромосом и генов. Этот набор остается диплоидным в соматических клетках и гаплоидным в половых. 8. Копуляция половых клеток с одинаковым гаплоидным набором обеспечивает образование в зиготе следующего поколения — диплоидного набора родительского типа. 9. Следовательно, воспроизведение родительских признаков в потомстве объясняется передачей ему соответствующих генов родителей.
Стройность этой концепции в общем способствовала укреплению представления о генах, как обособленных единицах наследственности. Вопрос был поставлен таким образом, что отношения между геном и признаком могут быть определены формулой «ген → признак», подобно, тому, как Вейсман говорил о детерминантах, как определителях признаков, или ранние менделисты — о факторах, ответственных за определенные признаки. Морган, как и ранние менделисты, рассматривал особь не как наследственно-целое, а как суммированное фенотипическое выражение обособленных единиц наследственности. Правда, между вейсманистской и морганистской концепцией генетического содержания организма имеются принципиальные различия. Морганизм пришел к существенно иному пониманию генной структуры соматических клеток. Вейсман принимал, что деление клеток зародыша идет путем качественно неравного распределения детерминантов, почему каждый орган получает особый набор детерминантов, представляющих данный орган и отличный от набора детерминантов в любом другом органе. Согласно же морганистской концепции, во всех органах все клетки имеют качественно однозначный набор генов (качественно равное, или эквационное, деление клеток). Концепция эта безусловно правильна.
Из сказанного выше видно, что представление об организме, как суммированной мозаике признаков, и генотипе, как сумме генов, все же не было устранено. Гены уподоблялись «бусам на нитке» (Морган), а хромосомы в целом трактовались как суммированное представительство «пучка» признаков.
Г. Взаимодействие генов. Однако, это представление, в сущности, уже к концу того же первого десятилетия XX века потеряло почву под ногами и морганизм отошел от него. Это было связано с дальнейшем развитием теории взаимодействия генов. По мере накопления сложных случаев расщепления, выяснилась необходимость связывать развитие каждого конкретного признака со многими генами. Такие гены, влияющие сообща и оказывающие на конкретный признак суммированное действие, были названы полимерными. Нильсон-Эле (Nilson-Ehle, 1909) обнаружил полимерию при скрещивании овсов с черными и белыми чешуями.
При скрещивании этих форм ожидалось моногибридное расщепление (3:1), в действительности же было получено отношение (15:1). Это было объяснено при помощи предположения, что за черную окраску чешуи ответственны две пары однозначных, или полимерных, генов. Подобные явления позднее были обнаружены в очень большом числе случаев, и полимерия оказалась весьма частым явлением. Сущность полимерии состоит в целостном действии различных генов на один и тот же признак. Это целостное действие генов носит при этом различный характер. В одних случаях различные гены воздействуют сообща на детали одной и той же части тела. Например, несколько генов участвует в развитии дикой окраски остевых волос мышей. Ген С обеспечивает появление пигмента вообще, ген В — чернокоричневого пигмента, Е — потемнение меха, ген А распределяет черный пигмент у основания волоса и желтый — на его вершине, Д — усиливает всю пигментацию волоса и т. п. В других случаях наблюдается полимерия однозначно действующих генов, когда разные гены обеспечивают развитие одного и того же признака, как это было сказано для черных чешуй овса.
С другой стороны, было предположено, что один и тот же ген воздействует на многие признаки. Например, принимается, что у мышей один и тот же ген влияет на окраску шерсти (вызывает желтую окраску), на плодовитость, на жировой обмен, на жизнеспособность зародышей и т. п. Такие гены называются плейотропными. В других случаях один и тот же ген воздействует на различные признаки, относящиеся к различным стадиям развития (плейотропизм во времени, Шмальгаузен, 1938). В этом случае один и тот же ген участвует как в окончательной моделировке данного органа, так и на поледовательных стадиях развития его.
Развитие этик идей показало, что в общем все признаки так или иначе связаны с любым геном, и все гены так или иначе воздействуют на любой признак, возникающий в ходе онтогенетического развития. Совершенно ясно, что эти представления возникли из усложненных случаев расщепления потомства. Мы уже видели, что при несоответствии действительно полученных числовых отношений потомства менделевскому отношению, в систему подставлялись дополнительные гены, которые позволяли свести любое числовое отношение потомков к менделевскому. Правомерность этого приема основана на следующем. Были, например, наблюдены следующие числовые отношения второго поколения при дигибридном скрещивании: 9:3:3:1 (меиделевское), 9:3:4, 9:7, 15:1, 12:3:1, 13:3. Легко констатировать, что во всех этих случаях сумма чисел равна 16. Следовательно, все указанные числовые отношения потомков возникли в результате комбинации четырех типов мужских и четырех типов женских гамет, т. е. соответствуют дигибридному расщеплению. Если это так, то для получения возможных шестнадцати комбинаций (4X4) необходимо в формулу гамет подставить столько предполагаемых генов, сколько нужно для сведения данного случая к менделевским отношениям (4X4=16).
Разберем еще один пример, показывающий способ обоснования представлений о полимерных генах.
Шелл (1914) скрестил растения одного вида пастушьей сумки Capsella bursapastoris с другими — С. Heegeri. У первого вида плоды треугольные, у второго — овальные. Во втором поколении произошло расщепление приближенно в отношении 15 треугольных к 1 овальному. Очевидно, вопрос идет о дигибридном расщеплении (15 + 1 = 16). Было прослежено третье поколение, чтобы проанализировать генетический состав F2. Оказалось, что в F2 растения с овальными плодами дают чистое потомство, а потомство растений с треугольными плодами вновь расщепляется на три группы: 1) растения с овальными плодами, 2) растения с треугольными и овальными плодами в отношении 3:1 и 3) те же типы в отношении 15:1. Если предположить, что по данным признакам имеется четыре типа гамет, то тогда описанное явление может быть уложено в шкалу дигибридного скрещивания (4X4=16=15+1). Было предположено, что треугольная форма плодов определяется двумя полимерными генами Т1 и Т2. Следовательно, гаметы их имеют формулу Т1Т2. Овальная форма связана тогда с рецессивными генами t1 и t2, и следовательно, их гаметы имеют формулу t1t2. Гибридная зигота и гибридное растение, полученное от скрещивания этих гамет, будет иметь формулу T1t1 T2t2. Скрестив такие два растения, мы получим гаметы: Т1Т2, T1t2 t1T2 и t1t2. На рисунке показаны результаты их скрещивания, дающие отношение 15:1 и являющиеся, очевидно, видоизмененным менделевским отношением 9:3:3:1. Таким образом, допущением существования полимерных генов Т1 и Т2, суммарно определяющих треугольную форму плодов, отношение 15:1 было уложено в решетку Пеннета.
Наследование треугольной и овальной формы стручков у пастушьей сумки (Из Рокицкого)
Как видим, представление о дополнительных генах сыграло роль рационального способа объяснения сложных случаев расщепления. Но вместе с тем это представление привело к развитию идеи о взаимодействии генов (полимерия, плейотропизм).
Д. Наследственное основание, как целостность. Идея взаимодействия генов привела генетику к представлениям, которые имели очень важные следствия. Стало ясным, что понимание генов, как функционально обособленных единиц наследственности — неправильно. Очевидно, хромосому нельзя понимать, как сумму генов. Представление о них, «как бусах на нитке», должно было пасть. Вместо него возникло другое представление о хромосоме, как целостной системе, в которой гены функционально связаны в некоторое единство («континиум»), в пределах которого изменение в одной части хромосомы влечет за собой изменение действия других частей ее.
Идея эта получила теоретическое обобщение в теории эффекта положения, предложенной Стертевантом (1925) и тесно связанной с представлением о линейном расположении генов в хромосоме. Согласно этой теории, при перестройках хромосом нужно предположить, что конкретные гены вступают в новые связи с другими генами, и так как они взаимодействуют друг с другом, то в новом положении их действие, их эффект становится другим. В новом положении гены обусловливают возникновение новых признаков.
Критика теории гена
К критике теории гена нужно подходить с известной осторожностью, так как при современном состоянии наших знаний мы не можем предложить какой-либо другой теории, столь же конкретно объясняющей нам большие группы явлений наследственности. Поэтому значительно правильнее в критике теории гена исходить из нее самой. Мы здесь ограничимся лишь теми сторонами критики теории гена, которые связаны с основными теоретическими принципами дарвинизма.
Мы видели, что путь развития морганизма привел его от представления об обособленных генах к необходимости понимания их в тесной функциональной взаимообусловленности. Хромосома стала восприниматься, как целостный, но качественно диференцированный вдоль её длины элемент клетки, любые отрезки которого связаны взаимодействиями. Был высказан взгляд, что изменения в одном участке хромосомы («в одном гене») распространяются вдоль хромосомы и что, следовательно, эти изменения выражены в протекании вдоль хромосомы каких-то биохимических и физиологических процессов. Мы видели также, что согласно теории эффекта положения, перекомбинацця (перемещение) генов приводит к изменению их действия. Казалось бы, эти представления должны были привести к идее развития генотипических структур. Если наследственное основание есть целостность, то, как всякая целостность, оно должно быть внутренне противоречивым и, следовательно, должно развиваться, в диалектико-материалистическом понимании термина «развитие». На деле морганизм рассматривает наследование не как процесс развития, не как форму движения, а как явление передачи потомку константных («готовых», по выражению Тимирязева) генов. Гены копулирующих гамет суммируются в зиготе и, следовательно, гены потомка — это суммированные гены родителей. Как видим, наследование понимается здесь не как процесс развития, а как процесс передачи константных генов от поколения к поколению. Мы не примем этого взгляда, исходя из положения, что процесс наследования нельзя свести к наследственности, и, напротив — он всегда является диалектически противоречивым единством наследственности и наследственной изменчивости, т. е. развитием генотипических структур. Правда, морганизм принимает, что отдельные гены мутируют. Мутирование генов рассматривается при этом, как редкое и спорадическое явление. Обычно указывается, что редкость мутаций отдельных генов «необходима эволюционной точки зрения» (Бельговский, Кирпичников и Прокофьева-Бельговская, 1940), так как если бы каждый ген изменялся в каждом поколении, то результаты творческого действия естественного отбора не закреплялись бы в поколениях. Значение этого соображения, в известной мере, ограничивается, однако, указаниями самих генетиков, что так как в каждой хромосоме имеются сотни генов, то общая мутационная изменчивость очень велика. У дрозофилы она достигает, по последним данным, 40% общего числа гамет. Фактически она, вероятно, больше, и следовательно геном, как целое, изменяется очень часто, тогда как результаты отбора этим не устраняются. Объясняется это тем, что отбор идет не по генам, а по признакам и что одни и те же признаки сплошь и рядом развиваются на основе разных геномов. Поэтому попытка использовать факты отбора, как аргумент в пользу теории необходимости постоянства генов не является бесспорным. В генетике, на почве излюбленной идеи постоянства генов, создались следующие представления о происхождении мутаций. Морганизм принимает, что мутации имеют троякое происхождение. Обычно различают: 1) Трансгенации — мутации «отдельно взятых» генов, т. е. изменения в определенных точечных локусах хромосомы; 2) хромосомальные аберрации — мутации, возникающие вследствие перестроек хромосом, сводимых к перекомбинации генов и их новому эффекту в новом положении (но без изменения самих генов); 3) мутации кариотипа (совокупность видимых во время кариокинеза, мейозиса и оплодотворения яйцевой клетки хромосом, их типичное для данного вида число, форма и т. п.). Мутации кариотипа выражаются, как правило, в изменении числа хромосом.
Примером кариотипической изменчивости могут служить явления полиплоидии и амфидиплоидии. Эти явления наблюдаются и в природе, но были изучены преимущественно экспериментально, в частности — действием на семена и прорастающие растения специфических веществ. Особенное распространение получил метод воздействия на прорастающие семена слабыми растворами колхицина (добывается из Colchicum autumnale). Метод этот был предложен Блексли и Айвери (1937).
Амфидиплоидия оказалась связанной с интенсификацией ряда признаков растений, увеличением размеров, значительным увеличением числа хлоропластов в зеленых частях растения, возникновением и повышением плодовитости (у стерильных гибридов), увеличением размера и веса семян и рядом других изменений. Самый процесс удвоения числа хромосом объясняется следующими причинами. Когда материнские хромосомы расщепляются, то нормально каждая продольная половина хромосомы оттягивается к противоположным полюсам клетки. Под влиянием колхицина, повидимому, изменяется вязкость плазмы (Кольцов, 1939), что препятствует образованию ахроматинового веретена и имеет следствием нерасхождение дочерних хромосом к полюсам клетки. Следовательно, их число так и остается удвоенным, а клетка становится амфидиплоидной.
Возник вопрос, какие же из названных изменений служат истинной причиной новых мутаций? Очевидно, только трансгенации, так как остальные изменения понимаются, как новые комбинации прежних генов. Такой постановкой вопроса генетика открыла широкие двери для проникновения в ее систему чисто метафизических представлений. Было, в частности указано, что нет возможности установить экспериментальные различия между мутациями, возникшими вследствие перестройки хромосом (перекомбинации генов) и мутациями, происшедшими вследствие изменения генов. Некоторые генетики сделали попытку свести все мутации к эффекту положения, т. е. к перекомбинациям константных генов. Вопрос об изменяемости генов становится, таким образом, делом личных взглядов. И действительно, одни генетики принимают изменяемость генов, а другие — отрицают ее. Подчеркнем, что это диаметрально различное отношение к вопросу строится на основе одной и той же теории гена.
Попробуем представить себе дарвиниста, который отрицал бы эволюцию видов. Такое положение невозможно, так как эволюция видов с необходимостью вытекает из теории естественного отбора. В теории же гена, изменяемость генов не вытекает из теории, как необходимое следствие ее. Именно в этом основной дефект морганистской теории гена. Не удивительно поэтому, что одни из генетиков, стоящие на почве этой теории, разделяют точку зрения дарвинизма, а другие приходят к антиэволюционным выводам. Ясно, что такое положение дискредитирует теорию гена. Очевидно, она должна быть построена так, чтобы изменяемость генов с необходимостью вытекала из нее, иначе возможны рецидивы антиэволюционных воззрений.
Такая постановка проблемы гена связана с отказом от чисто морфологического анализа явлений наследования, еще недавно господствовавшего в современной генетике, которая оперирует, по преимуществу, с морфологическим анализом хромосом и признаков, их числом, формой, соотношениями и морфологическими изменениями. Этот по преимуществу морфологический аспект на закономерности и причины наследственности вскрывает лишь форму процесса, но не дает причинного объяснения содержания его, приобретая именно поэтому формальный характер. Неудивительно поэтому, что многие генетики возражали и возражают против по преимуществу морфологического аспекта и склоняются к необходимости физиологического и биохимического понимания процессов наследования. На этот путь в свое время стал, например, В. Ружичка (1909), который считал, что наследственность, как свойство, основана «на особом химическом строении и на обусловленных последним, при известных условиях, процессах обмена». В новейшее время Гольдшмидт (1938) предложил теорию «физиологической генетики», в которой хромосомы рассматриваются как целостные генетико-физиологические реактивные системы — гигантские цепные молекулы, а гены — лишь как ее радикалы, связанные в своем действии с другими радикалами целостной системы хромосомы. Эти представления безусловно интересны и в своей основе были бы правильны, если бы Гольдшмидт не придал им чисто механистического характера, сведя изменяемость генов (радикалов) к количественным изменениям, а именно — что изменение действия гена является следствием изменений числа молекул, входящих в его состав. Развитие генетической структуры сводится здесь к росту (увеличению и уменьшению), а не рассматривается, как качественные превращения. Значительно правильнее поэтому, оставаясь на почве физиологического и биохимического подходов к явлениям наследственности, следовать за общими идеями К. А. Тимирязева и И. В. Мичурина, основные принципы! которых могут быть выражены в следующих положениях.
1. Наследование есть процесс развития, всегда связанный с условиями, в которых оно протекает.
2. Процесс наследования теснейшим образом связан с физиологическими процессами, протекающими в клетке, в свою очередь, обусловленными теми связями, которые всегда имеются между клеткой и внешней, по отношению к ней, средой.
3. Процесс наследования не может быть сведен к передаче «готовых» (по выражению Тимирязева) корпускул наследственного вещества — генов (сравни п. 1).
Изложенные выше принципы мы положим в основу дарвинистской интерпретации учения о наследственности, так как в системе дарвинизма наследственность нас интересует не сама по себе, а как процесс наследования, всегда имеющий две противоречивые, диалектически связанные стороны: наследственность, как стойкость структур и реакций клетки, и наследственную изменчивость, как процесс известного сдвига этих последних. Мы примем, что наследование есть процесс индивидуализированного (т. е. индивидуально измененного) воспроизведения потомком наследственных свойств родителей.
Чтобы убедиться в правомерности таких представлений, рассмотрим: 1) что такое наследственность и 2) что такое наследственные изменения.
Что такое наследственность?
Успехи, сделанные современной биохимией, позволяют утверждать, что способность к наследственной передаче определенных качеств от родителей к потомству определяется, в первую очередь, известной стойкостью биохимической структуры клетки. С другой стороны, следует помнить вполне оправдавшую себя формулировку Энгельса, что жизнь есть особая форма существования белковых тел. Белки же отличаются огромным многообразием. Системы их, как правило, весьма подвижны, лабильны и изменчивы. Следовательно, если наследственность есть известная стойкость биохимических структур клетки, то спрашивается: каковы же причины этой стойкости изменчивых и многообразных белковых тел? В сущности, в первую очередь, в этом и состоит проблема наследственности.
Клетка является сложной и внутренне диференцированной системой, органоиды которой специализованы и связаны между собою тесными биохимическими и физиологическими взаимозависимостями. Внутриклеточная диференциация клетки выражается, прежде всего, в разделении ее на цитоплазму и ядро. Обе эти части клетки различны как по своей морфологической и биохимической структуре, так и по функциям. В основе характеристики цитоплазмы лежит тот факт, что она непосредственно связана с внешней, по отношению к клетке, средой. Ядро, напротив, связано с ней посредственно, через цитоплазму. Этот факт имеет огромное значение, так как он доказывает, что материал ядра строится не непосредственно за счет внешней среды, а за счет материала цитоплазмы, которая имеет значение энергетического посредника между внешней средой и ядром. Уже наличие таких окольных связей с внешней средой указывает, что ядро должно обладать более стойкой биохимической структурой. И действительно, ассимилируя поступающие извне продукты, цитоплазма подвергает их биохимической обработке. Содержанием этих сложных биохимических и физиологических процессов является усвоение поступающих в цитоплазму веществ, в ходе которого «внешние» вещества качественно изменяются и превращаются в составные биохимические вещества самой цитоплазмы. В этом смысле можно сказать, что под влиянием обмена «внешнее» становится «внутренним». Однако, если это так, то отсюда понятно, что превращение «внешнего» во «внутреннее» означает известную нивеллировку внешних воздействий. Нивеллирующая роль цитоплазмы, обладающей конкретной биохимической структурой и связанными с последней типичными (для конкретного генома) формами обменных процессов, и есть одна из причин стойкости самой биохимической структуры.
С другой стороны, ядро, в свою очередь, воздействует на биохимию цитоплазмы и на обменные процессы в ней. Факты показывают при этом, что ядро участвует в этих процессах избирательно, воспринимая совершенно определенные продукты цитоплазмы. Поэтому биохимически ядро отличается от цитоплазмы. Оно содержит особые ядерные белки — нуклеопротеиды, характерные именно для ядра, образующиеся, однако, несомненно за счет цитоплазмы. Избирательные свойства ядра, воспринимаются как совершенно определенные продукты и являются другой причиной стойкости биохимических структур клетки. В конечном же счете, нивелирующая работа цитоплазмы и избирательные свойства ядра имеют своим следствием стойкость именно ядерной биохимической структуры. Если, однако, ядерная биохимическая структура ядра стала, по указанным выше причинам, стойкой, то отсюда понятно, что под влиянием ядра становится относительно стойкой вся биохимическая система клетки. Эта стойкость ее и обеспечивает стойкость реакций клетки, как целостной системы, т. е. — стойкость нормы реакций на внешние воздействия. Но ведь именно норма реакции и есть основа характеристики любого конкретного наследственного основания. Таким образом, отвечая на вопрос, что такое наследственность, в чем ее сущность, мы можем, прежде всего, констатировать, что наследственность есть относительная стойкость биохимической структуры и реакций клетки, и в частности ее хромосомного аппарата.
Нельзя при этом упускать из виду и морфологическое строение хромосом, как фактор, повышающий стойкость их биохимической структуры, — вопрос, который мы оставим в стороне. Из сказанного выше понятно, почему именно ядро становится ведущим генетическим аппаратом клетки. Очевидно, что ядро как бы аккумулирует в себе, в своей структуре, основную генетическую характеристику клетки. При этом нужно, конечно, помнить, что эта роль ядра возникает в его биохимических связях с цитоплазмой. Поэтому мы должны признать, что, строго говоря, генетическим аппаратом является вся клетка, как целое, и что ядро превратилось в специальный аппарат, который позволяет относительно фиксировать сложившиеся генетические свойства клетки. Однако, на сказанном проблема причин стойкости наследственных структур не заканчивается.
Одной из важнейших жизненных функций клетки является ее деление, так как именно этим путем обеспечивается передача её свойств дочерним клеткам. Элементарные факты показывают, что деление клетки (кариокинез) идет по типу эквационного деления, т. е. обе дочерние клетки получают качественно и количественно равный ядерный материал. Исторически) эквационное деление было достигнуто путем диференцировки наиболее стойких биохимических структур ядра на отдельные хромосомы. Факты показывают, что «поведение» хромосом — целесообразно. Как правило, они имеются в четном числе, их сочетания при кариокинезе, мейозисе и оплодотворении — арифметически точны. В самом деле, редукционное деление (мейозис) идет, как правило, по формуле 2n:2 = n, а оплодотворение по формуле n + n = 2n. Этот поразительный факт явственно показывает, что хромосомы являются специальным генетическим приспособлением клетки, обеспечивающим равномерное распределение биохимически наиболее стойкой, а потому и наследственной субстанции. Таким образом, хромосомы являются генетическим приспособлением клетки, ее приспособительным передаточным генетическим аппаратом, обеспечивающим наследование.
Этот факт приводит к выводу, что поразительное приспособительное «поведение» хромосом не было дано заранее. Напротив, оно исторически выработалось еще у древних одноклеточных организмов, под влиянием естественного отбора, через уничтожение обладателей менее гармоничных хромосомных систем и, следовательно, сохранение более гармоничной организации хромосомного аппарата, обеспечивающего правильное распределение наследственных качеств между дочерними клетками и гармоничность реакций развивающегося организма (неправильные, так называемые «несбалансированные» наборы хромосом, как правило, влекут за собой дезинтеграцию геномов, а вследствие этого — нормы реакции и нарушение хода индивидуального развития).
Таким образом, ядро и хромосомы приобрели ведущее значение в передаче наследственных свойств, как по биохимическим, так и по историческим причинам.
При этом, однако, следует подчеркнуть, что все же, как уже говорилось, генетико-физиологическим аппаратом остается клетка, как целое.
Именно поэтому наследственные качества клетки, как целого, как бы «не умещаются» в ядре. В самом деле, в генетике сложилось представление, что кроме хромосомных генов существуют плазматические гены. Поэтому на ряду с понятием генома, как целостной системы хромосомных генов, было предложено понятие плазмона (Wettstein, 1928), как системы плазматических генов. Ряд фактов, на которых мы не будем останавливаться, привел некоторых генетиков к мысли, что как правило ядерные (хромосомные) гены определяют частные признаки, а плазматические — более общие. Эти представления следует считать дуалистическими и механистическими, так как они приводят к мысли о существовании в клетке каких-то двух независимых систем корпускул наследственного вещества. В действительности факты плазматической наследственности говорят именно в пользу представления, что клетка является целостным генетическим аппаратом. Хромосомы должны, следовательно, пониматься не как вместилище «отдельно взятых» независимых генов, а как качественно диференцированные сложные белковые системы, фиксирующие, в ходе обменных процессов между ядром и плазмой, определенную биохимическую структуру, а следовательно, и генетические функции целостной клетки. Следовательно, генный состав хромосом отображает всегда не только генетическое состояние самих хромосом, но и генетическое состояние всей клетки, как целого. Ввиду сказанного, нет никакой надобности различать «геномы» и «плазмоны», и в дальнейшем мы будем говорить только о клеточном геноме, как биохимически и функционально целостной генетико-физиологической системе клетки.
Мы можем резюмировать разобранные представления о наследственности.
1. Наследственность есть исторически и биохимически сложившаяся относительная стойкость биохимической структуры и реакций клетки, и в особенности ее хромосомного аппарата.
2. Любая хромосома не есть, однако, агрегат «отдельно взятых» автономных генов, но качественно диференцированная сложная белковая система, биохимическая структура которой, а следовательно, и ее генетические качества всегда являются зафиксированным результатом генетико-физиологического состояния целостной клетки.
3. Вследствие возникшей, по биохимическим причинам, стойкости структуры хромосом, они исторически превратились в постоянный приспособительный передаточный генетический аппарат, обеспечивающий равномерное распре деление наследственных качеств материнской клетки, как целого, между дочерними клетками.
Что такое наследственные изменения?
Стойкость биохимических структур хромосом и клетки, как целого, не следует, однако, преувеличивать. Нельзя упускать из виду неисчерпаемого многообразия белковых структур и их потенциальной подвижности. Биохимическая структура хромосом не есть система равновесия, абсолютного покоя. Напротив, самое существование ее обусловлено ее развитием, изменениями системы, постоянным взаимодействием с остальными частями ядра и цитоплазмы. Нельзя допустить, как это иногда делается, что ядро влияет на функциональное состояние цитоплазмы и структуру ее, а цитоплазма влияет на функциональное состояние ядра, но не влияет на структуру ядра. Весь вопрос в том, о какой структуре идет речь. Морфологическая структура может быть не изменена. В биохимической структуре хромосом происходят, однако, сдвиги, обусловливающие и генетические изменения. Вопрос лишь в том, когда и где именно последние происходят.
Чтобы ответить на этот вопрос, мы должны еще раз остановиться на понятии генома. Здесь нужно подчеркнуть, что геном означает именно целостную материальную основу генетического аппарата, его скоррелированность. Именно в этом смысле следует говорить о геномных корреляциях. Однако, целостность никогда не бывает заранее дана. Напротив, она становится и развивается. Положение это четко показано Шмальгаузеном (1938) для индивидуального и исторического развития организма, — его формообразования. Это целиком остается справедливым и для генома; геном не дан заранее, а становится и развивается. Однако, это означает, что в известные моменты жизни клетки он разрушается и вновь строится. Именно в эти переломные моменты и возникают наследственные изменения. Мы знаем, в основном, два таких момента: 1) период созревания половых клеток и 2) момент оплодотворения.
Многочисленные данные показывают, что мутации происходят большей частью в половых клетках. Но ведь именно в половых клетках возникают процессы, ведущие к «ломке» того генома, который установился в системе и в клетках данного организма. Это — процессы созревания половых клеток и в частности явления мейозиса (редукционного деления), приводящего к возникновению гаплоидного набора хромосом. С точки зрения морфолога-цитогенетика, гаплоидный набор хромосом есть уменьшение (вдвое) диплоидного набора. Диплоидный же набор, возникающий в оплодотворенном яйце, есть увеличение (вдвое) гаплоидного набора. Возникает весьма плоская концепция, сводящая развитие гамет и зиготы к уменьшению и увеличению, — концепция, опровергнутая В. И. Лениным. Созревание гамет и развитие зиготы, в свете биохимического и физиологического подходов к наследственности, есть, конечно, процесс развития в диалектико-материалистическом понимании его. Мутации возникают вследствие тех физиологических и биохимических процессов, которые разыгрываются в зреющих гаметах. С этой стороны очень интересны данные, которые показывают (Берман, 1939), что под влиянием рентгена частота мутаций в незрелых половых клетках ниже, чем в зрелой сперме, и что она (частота) различна на разных стадиях спермиогенеза. Это означает, что в процессе созревания происходят наследственные изменения. Берман пришел к выводу, что «различия физико-химического состояния половой клетки на разных стадиях спермиогенеза играют, повидимому, весьма важную (а может быть и решающую) роль в наблюдаемых различиях частоты мутирования». Берман полагает, что мутирование есть следствие изменения физиологического состояния половой клетки и физико-химического состояния самого гена. Это мнение далеко не одиноко. Напротив, многочисленные данные целого ряда исследователей показывают, что различные агенты — температура, химические факторы, состав пищи и т. п. — влияют на мутирование, и что за процесс мутирования ответственны физиологические и биохимические изменения, протекающие в среде, непосредственно окружающей гены, т. е. в самой среде ядра (Керкис, 1939, 1940).
Итак, в основе наследственных изменений лежат структурные биохимические изменения хромосом, возникающие под влиянием физиологических (обменных) процессов в половой клетке, в ее связях с внешними, по отношению к ней, факторами среды, и притом, как правило, в переломные (критические) моменты — в период созревания гамет. В самом деле, гамета, с ее гаплоидным геномом, не адекватна любой соматической клетке с диплоидным набором. Ее реакции — иные. Поэтому иными оказываются и отношения зреющей гаметы к внешним факторам. Отсюда известная перестройка (развитие) генотипических структур. Таким образом, мутирование не есть спорадическое явление, а процесс развития, «конечным» итогом которого являются мутации) в их гено — и фенотипическом выражении. При такой постановке вопроса естественно возникает представление, что каждое поколение способно мутировать. Это положение не чуждо современной генетике. Если мы проследим историю учения о мутациях, мы увидим, что представления о частоте мутационного процесса изменялись по мере углубления знаний о данном биологическом объекте. Уже исследования Баура (1924) над львиным зевом показали, что число мутаций очень велико и что большинство из них выражается в мелких изменениях физиологического характера. В диких популяциях дрозофилы обнаружено огромное количество мелких физиологических и морфологических мутаций (Балкашина и Ромашев, 1935; Дубинин, 1940). В настоящее время принимается, что мутирует около 40% гамет. Керкис (1940) указывает, что «огромное большинство возникающих мутаций затрагивает мелкие, внешне почти не проявляющиеся физиологические признаки организма, вследствие чего они ускользают от обычных наблюдений». Это значит, что вопрос идет о таких мутациях, которые не изменяют или почти не изменяют нормы реакции организма, т. е. почти не выходят за прежние пределы ее порогового значения. Такие мутации вследствие этого становятся латентными (скрытыми). Именно эти мелкие наследственные сдвиги, идущие из поколения в поколение, и составляют основу мутационного процесса. Нужно поэтому согласиться с Навашиным и Герасимовой (1935), которые высказываются против представлений о спорадичности мутаций и указывают, что развитие мутационного процесса идет с такой же необходимостью, как всякое развитие.
Биохимический и физиологический подходы к явлениям наследственной изменчивости нашли свое подтверждение и в своеобразных явлениях «старения» семян, описанных М. С. Навашиным.
Исследуя мутационный процесс, М. Навашин (1933) обнаружил, что в «стареющих» семенах скерды (Crepis tectorum) наблюдается повышение частоты наследственных изменений. Было высказано предположение, что последние возникают вследствие изменений обменного порядка. Чтобы подтвердить сказанное, было допущено, что под влиянием внешних факторов, усиливающих процессы физиологических и физико-химических превращений в семенах, должно наблюдаться и усиление наследственных изменений. Семена скерды были помещены в термостат при температуре 50—60°. Двадцатидневное действие этой температуры дало повышение частоты наследственных изменений, равное, по своему эффекту, 6—7-летнему хранению семян, не подвергавшихся никакому воздействию (Шкварников и Навашин, 1935). Таким образом, простое повышение температуры ускорило протекание процессов наследственных изменений. Однако, они протекали и без этого вмешательства, только медленнее. Эти отношения с очевидностью указывают на то, что за процесс наследственных изменений ответственны биохимические и физиологические превращения в семенах, связанные конечно с обменными процессами, а следовательно и с отношениями внешней средой.
Наблюдения Навашина и Шкварникова имеют очень большую методологическую ценность, так как они показывают, что в генетикофизиологической системе действительно протекают конкретные процессы развития генотипической структуры. Возможность ускорения этих процессов является доказательством их существования.
П. М. Жуковский (1944) указывает, что в 1928 году им были скрещены злаки Гайнальдия (Haynaldia villosa) и Triticum dicoccum var. farrum. Часть полученных гибридных семян была высеяна лишь в 1938 году, т. е. спустя 10 лет. Все полученные растения оказались амфидиплоидными. По Жуковскому, амфодиплоидия возникла здесь вследствие «старения» при долголетнем хранении семян, вызвавшем наследственные изменения.
Эти случаи явственно указывают на то, что в «стареющих» семенах, под влиянием обменных процессов имели место физиологические сдвиги, приведшие к мутациям.
В свете изложенных представлений возможно соответственным образом подойти и к явлениям хромосомных перестроек. Физиологический и биохимический подходы к явлениям наследственности привели более вдумчивых генетиков к выводу, что представления об отдельных генах снимаются развитием самой генетики (Керкис, 1940). Это означает, что становится спорным чисто корпускулярное представление о гене. «Отдельно взятых» генов — нет. Гены же в их тесных физиологических и биохимических связях, т. е. как выражение качественной диференцировки хромосом — реальное явление. Наличие связей между генами и их несомненная биохимическая природа заставляют считать, что изменения в одном участке хромосомы неизбежно связаны с изменениями в других частях ее. Изменения в «одном гене» (в известной части хромосомы) влечет за собою какие-то процессы вдоль хромосомы. Эти представления согласуются с новейшими данными о видимой морфологической структуре хромосом. Исследования некоторых ученых показали, что хромосомы имеют сложную биохимическую структуру (Гейц, 1933; Прокофьева-Бельговская, 1939, 1940; Дарлингтон, 1942 и др.) и состоят из биохимически различных, «эухроматических» и «гетерохроматических» участков, отличающихся по красочным реакциям. Оказалось, что «гетерохроматические» участки относительно легко изменяются под влиянием различных физиологических агентов и в свою очередь изменяют «эухроматические» участки хромосом. Далее, было выяснено, что изменения в «эухроматических» участках связаны с изменениями признаков. Наиболее явственные генные изменения (мутации) происходят в участках, непосредственно прилегающих к гетерохроматическим районам (Прокофьева-Бельговская, 1939). Таким образом, «гетерохроматические» участки являются как бы проводниками внешних физиологических воздействий, и через их посредство изменения распространяются вдоль хромосомы. Эти данные позволяют защищать представление, что и хромосомные перестройки не являются элементарной перекомбинацией константных генов. Следует задаться вопросом о причинах возникновения самих перестроек. В свете биохимического подхода может быть только один ответ — вследствие биохимического взаимодействия между генами, между хромосомами и между ними и цитоплазмой. Хромосомные перестройки, следовательно, едва ли являются перетасовкой абсолютно константных генов и должны пониматься, как выражение тонкой биохимической перестройки хромосомы под влиянием физиологических и биохимических сдвигов в клетке. Поэтому эффект нового положения гена есть на самом деле эффект относительно измененного гена. Новые биохимические связи между участками хромосомы означают и относительно новые биохимические качества. Хромосомные перестройки (независимо от их величины, вплоть до «точечных» изменений и изменений, лежащих за пределами микроскопической видимости) означают тонкие биохимические сдвиги — относительное обновление генной диференцировки хромосомы, а не перекомбинацию константных генов.
Возникает вопрос: какова частота этого процесса относительной генной передиференцировки хромосом? В суждении об этом вопросе правильнее всего исходить из частоты мутаций и их величины. У дрозофилы, как было выше указано, частота мутаций равна 40%. Число это значительно больше предполагавшегося раньше, и увеличилось оно преимущественно за счет мелких мутаций. Нет никаких оснований к тому, чтобы отказаться от предположения, что в дальнейшем выяснится значительно большая частота мутаций. Если принять, что в приближении частота мутаций определяется 50%, то это означает, что из каждых двух гамет одна мутирует. Уже эти отношения позволяют высказать предположение, что по крайней мере в 50% генная структура хромосом образуется относительно заново. С методологической точки зрения, перестройки хромосом становятся тогда выражением развития, а не перекомбинации константных генов. Однако, если учесть, что представления о частоте мутационного процесса изменялись за счет возрастающего обнаружения мелких мутаций, то не будет ошибкой принять, что фактически каждая гамета способна мутировать, хотя эти мутации и носят латентный характер, не выходя за пределы порогового значения прежней нормы реакции и не влияя, следовательно (эмпирически заметно), на формообразование организма. При такой постановке вопроса наследование становится формой развития, а изменяемость генов — необходимым теоретическим выводом. Поэтому правильнее говорить не о частоте мутаций, я об интенсивности их — от крупных до латентных.
Эти представления не противоречат понятию о наследственности, ее фактам и закономерностям. Они только подчеркивают, что наследственность, как стойкость структур, не есть абсолютный покой системы, ее «равновесие», что она всегда связана с изменчивостью, что, наконец, процесс наследования всегда включает наследственность (структурную стойкость наследственной материальной субстанции) и наследственную изменчивость, как две стороны одного и того же целостного процесса.
В перспективе мы можем допустить, что в каждом поколении генная структура диференцируется относительно заново. Этот взгляд допускается исследователями в области химии протоплазмы, и мы его примем. Разумеется, сказанное выше не означает, что всякий раз образуются совершенно новые гены. Новая генная диференцировка преемственно связана с родительской и идет по приближенно той же схеме, но всегда индивидуализированно. Повторение (рекапитуляция) генной структуры не есть абсолютное повторение, оно не есть механическая передача «готовых» генов, а форма развития генотипических структур.
Рассмотрев один источник наследственных изменений, перейдем к анализу другого источника их.
Геномогенез
У форм, размножающихся партеногенетически, мутации в зреющем яйце являются основным ближайшим источником развития наследственного основания. У организмов, развивающихся в результате копуляции гамет, последние «суммируются» и возникает зигота с диплоидным набором хромосом. Процесс этот происходит во время оплодотворения яйца. Образование зиготы, как генетической целостности, протекает быстро, но сопровождается коренными, глубокими биохимическим и физиологическим сдвигами в системе оплодотворенного яйца. Под влиянием оплодотворения происходит так назваемая активация яйца, конечным этапом которой является установление первой митотической фигуры и последующее дробление оплодотворенного яйца. Установление первой митотической фигуры означает, что «суммированные» геномы гамет приобрели качество организованного целостного генома новой зиготы. Однако, геном зиготы нельзя рассматривать, как простую сумму хромосомных наборов гамет. Геном потомка — не сумма, а новое качество, продукт развития нового наследственного основания. Справедливость этого взгляда вытекает, прежде всего, из того факта, что геном есть, как это уже отметалось, — целостность и что гены связаны в систему соответственными геномными корреляциями. Совершенно подобно тому, как целостность организма не заранее дана, а становится в ходе онтогенеза, так и геном, как скорелированная целостность, с очевидностью не дан заранее, а становится и развивается. Следовательно, каждый новый геном есть продукт развития, возникший на почве комбинирования (а не простого «суммирования») геномов копулирующих гамет.
Процесс организации генома новой зиготы мы и будем называть геномогенезом. Образование нового генома происходит на почве биохимических и физиологических сдвигов в оплодотворенном яйце. Соединение мужской и женской гамет есть, следовательно, процесс, в ходе которого возникает новый, диплоидный геном. Замечательно, что отцовские и материнские хромосомы сохраняют при этом свою морфологическую индивидуальность и свои генетические качества. Тем не менее остается безусловно ошибочным представление, что их генетическая природа в новой зиготе абсолютно адекватна родительской. Среда яйцевой плазмы не может не оказать на хромосомы известного структурного воздействия, которое тем сильнее, чем более чужеродны этой среде погруженные в нее хромосомы. В условиях отдаленных скрещиваний известны случаи разрушения отцовских хромосом, их элиминация. Но ведь чужеродность имеет свои степени. Полной адекватности, биохимической равнозначности отцовской и материнской плазмы нет. В какой-то мере любые две гаметы биохимически различны, хотя бы и в малой степени.
Поэтому и при родственных скрещиваниях отцовские хромосомы, погруженные в плазму яйца, должны испытывать хотя бы и очень тонкую биохимическую перестройку, не нарушающую их основной генной структуры, но все же несколько изменяющую ее. Сказанное справедливо. вероятно, и для материнских хромосом, так как плазма активизированного оплодотворением яйца биохимически не абсолютно равнозначна плазме неоплодотворенного яйца. Таким образом, генный комплекс зиготы подвергается тонким перестройкам, приводящим к установлению новой скоррелированной генной системы — генома новой зиготы. Эта представления не противоречат факту расщепления потомства в последующих поколениях. Однако, было бы методологически правильнее принять, что на фоне расщепления происходят тонкие генотипические сдвиги. Поэтому, например, формулу расщепления Aa x Aa = AA + Aa + aA + aa было бы правильнее изображать так: Aa x aA = AAn + Aan + aAn + aan, где n означает степень генотипического сдвига, т. е. степень геномогенеза.
Конечно, в случае близких (родственных) скрещиваний геномогенез отличается малой интенсивностью. Тем не менее, мы имеем право допустить его существование. Копуляция двух гамет приводит к возникновению комбинации, следствием которой будет комбинативная изменчивость, которую и можно трактовать как продукт геномогенеза, т. е. развития генома новой зиготы. Под комбинативной изменчивостью мы будем, следовательно, понимать не суммарную комбинацию двух гаплоидных геномов соединившихся гамет, а результат этого суммирования — интеграцию качественно относительно новых геномов, продуктов геномогенеза.
Таким образом, излагаемое понимание комбинативной изменчивости существенно отличается от проводимого в курсах генетики, где под комбинативной изменчивостью понимается только или по преимуществу сама комбинация, обычно расщепляющаяся в F2. В таком понимании самый термин «комбинативная изменчивость» (напомним, что изменчивость есть процесс изменений, а не измененные признаки) теряет свой истинный смысл, так как если последняя есть только расщепляющаяся комбинация, то о какой же подлинной изменчивости может идти речь?
Доказательством сказанного выше могут служить данные Сахарова и Магржиковской (1941, 1942), установивших, что при скрещивании дрозофил двух линий, но одного и того же вида (Drosophila melanogaster), частота мутаций в поколении, полученном от скрещивания, возрастает. Авторы указывают, что до их исследования было известно, что возрастание частоты мутаций наблюдается при гибридизации, т. е. скрещивании форм, относящихся к разным видам. Ими же установлен такой же факт в отношении скрещиваний в пределах одного вида. Усиление мутационного процесса есть прямое следствие скрещивания. Ясно, что последнее привело к известной дезинтеграции родительских геномов и к интеграции новых геномов, т. е. имело своим следствием геномогенез, который и выражен в усилении мутационного процесса в гаметах потомков. В практике сельского хозяйства пользуются подобными же скрещиваниями для разрушения («ломки») родительских геномов и создания новых, нерасщепляющихся геномов. Это создание новых геномов и есть геномогенез.
Наиболее явственно, однако, геномогенез наблюдается при отдаленных скрещиваниях, когда соединяются гаметы различных форм. Интенсивность геномогенеза в известных пределах прямо пропорциональна возрастанию различий между гаметами. Поэтому при отдаленных скрещиваниях геномы родителей подвергаются особенно интенсивной дезинтеграции, а гибридное потомство очень часто дает интенсификацию мутаций. Этот факт был доказан рядом авторов, скрещивавших разные виды, например — львиного зева (Баур, 1933), табака (Дончо Костов, 1935), хлопка (Харланд, 1937), дрозофилы (Бельговский, 1937). Мутации в гаметах гибридов являются здесь прямым следствием «ломки» родительских геномов и возникновения новых геномов.
В ряде случаев возникновение новых геномов (геномогенез) выражается в формировании новых нерасщепляющихся форм, полученных в результате гибридизации. Такие гибридогенные формы необычайно наглядно демонстрируют процесс геномогенеза. Таковы: гибридогенные нерасщепляющиеся формы, полученные через скрещивание двух видов пикульника, двух видов тимофеевки, малины и ежевики, тёрна и алычи, двух видов табака, пырея и пшеницы и т. д. (Дубинин, 1940). Во всех этих случаях совершенно ясно, что скрещивание привело к перестройке геномов и возникновению новых геномов, т. е. — процессам геномогенеза. Мы отмечаем здесь, что содержанием геномогенеза является развитие, возникшее на почве противоречий в комбинированном геноме, и притом — биохимического и физиологического порядка (разумеется, в связях соответственных генетических систем с факторами внешней среды). Всё сказанное выше позволяет считать, что геномогенез — реальное явление и что именно в нём содержание комбинативной изменчивости. Только в таком понимании последняя сохраняет свой истинный смысл — не простого суммирования хромосом, а процесса развития наследственного основания, т. е. процесса качественных сдвигов в нем. Поэтому следует согласиться с Навашиным (1941), что геном гибридной зиготы есть не суммарная комбинация, а новое качество. Можно допустить следующие отношения между мутациями и геномогенезом. Генетические противоречия между копулирующими гаметами являются источником геномогенеза, а новый геном становится источником новых мутаций в гаметах потомка.
Изложенная здесь гипотеза геномогенеза подчеркивает, что наследственность и изменчивость суть две противоречивые стороны единого диалектического процесса наследования.
Таким образом, геномогенез мы здесь трактуем, как процесс наследования, который приводит к интеграции нового целостного генома. Мы рассматриваем процесс геномогенеза, как форму развития, в ходе которого возникает относительно новая генная диференцировка зиготы. Так как зигота по своей биохимической и физиологической характеристике близка или даже приближенно идентична геномам родительской пары, то и генная диференцировка зиготы потомка протекает гомологично генной диференцировке исходных гамет, т. е. происходит рекапитуляция той же генной структуры, всегда, однако, относительно индивидуализирование. Разумеется, темпы этих процессов невелики. Геномогенез идет (как и мутирование) в форме мелких, скрытых незаметных изменений. Следовательно, как правило, наследственные изменения не влекут за собою резких генетических сдвигов. Если бы это было не так, мы имели бы дело с вечно изменяющимися рядами форм. Этого, однако, нет. Геномогенез приводит только к росту диалектических противоречий в целостном геноме. Геномы потомков в приближении такие же, как у родителей. Поэтому они сходны между собою. Однако, они не тождественны, так как их геномы индивидуализированны. Подобная трактовка процесса наследования (геномогенеза) необходимо вытекает из диалектико-материалистического понимания процессов. Целостность генома основана на диалектически связанных противоречиях, — источниках спонтанного развития. В системе генома этими противоречиями, в их связях с внешними факторами, являются стойкость (наследственность) и лабильность (изменчивость) белковых систем. Поэтому в системе генома стойкость и лабильность всегда сосуществуют, как стороны диалектического целого. Именно поэтому нет наследственности без изменчивости ни в одном последовательном поколении. Поэтому из поколения в поколение происходит не «передача» и не суммирование геномов гамет, а развитие генома или геномогенез.
К сказанному выше нужно прибавить, что сходство потомства с родителями покоится не только на сходстве геномов. Нужно четко различать преемственность генетического основания организмов (генетическая преемственность) и преемственность признаков (фенотипическая преемственность). Одни и те же признаки могут сохраняться на протяжении огромных промежутков времени и притом на различной генетической основе.
Геномогенез и онтогенетическое формообразование
Обратимся теперь, в кратких чертах, к характеристике отношений между геномогенезом и морфогенезом. Развитие нового генома морфологически выражается в образовании диплоидного набора хромосом, — конкретного кариотипа и в установлении первой митотической фигуры. Вслед затем, при наличии определенных внешних условий, начинается дробление яйца, которое (опять-таки при наличии определенных условий) сохраняет всегда типичные для каждого биологического объекта формы. Можно считать, что геном действует на дробление яйца в момент своего образования, так сказать, in statu nascendi и что именно под формативным воздействием генома, как целого, а следовательно и плазмы зиготы, и начинается типичный для данного биологического объекта процесс дробления. Именно в этом и выражается непосредственное действие генов, которые влияют, как первичный формативный фактор. Объясняется это тем, что гены продуцируют особые вещества — геногормоны. Однако, в дальнейшем процесс усложняется, так как, по мере дробления, клетки и ткани зародыша подвергаются все большей диференцировке. Возникает взаимно-воздействующая и взаимозависимая система, основанная на противоречивых связях между частями зародыша. При этом развивающиеся части тела оказывают взаимное формативное воздействие, обусловленное влиянием индуцирующих или формативных веществ, продуцируемых самими органами. Диференцировка частей зародыша приводит в этих связях к тому, что отношения между индуцирующим, формативным действием генов и развивающимися частями организма принимают иной характер. Непосредственным индуктором формообразования становятся продуцируемые тканями формативные вещества, которые и обусловливают возникновение сложных морфогенетических корреляций и зависимых перестроек. Например, мы уже знаем, что образование хрусталика зависит от формативного воздействия глазного бокала. Это, конечно, не означает, что действие генов прекратилось. Напротив, оно в сущности никогда не прекращается, но состоит в том, что гены определяют норму реакции клеточного материала, за счет которого образуется хрусталик под влиянием формативных веществ бокала. Другими словами, образование за счет клеток эктодермы именно хрусталика определяется не самим индуцирующим фактором (продуцированным бокалом), а реагирующим клеточным материалом. Такая постановка вопроса оказалась приложимой ко всем формообразовательным процессам, в частности и к процессу образования осевых органов (нервная трубка и хорда). Следовательно, именно внутренние свойства реагирующего клеточного материала определяют специфику его реакции. Однако, можно думать, что вопрос не сводим только к клеточному геному, т. е. к норме реакции клеток. Реакции клеток, конечно, имеются, однако, реакции тканей и частей организма, как целого, едва ли сводимы к реакциям клеточного генома. Это видно из того, что реакции одной и той же ткани на формативные вещества не остаются идентичными на все время онтогенеза. Включение в ход развития сложнейших морфогенетических и эргонтических корреляций делает взаимоотношения между геномом и морфогенезом в целом весьма сложными. Однако все же геном остается той общей организационной основой, вокруг которой возникают сложнейшие и далеко еще не выясненные процессы формообразования. В курсе дарвинизма для нас важно, однако, не обсуждение этих процессов (см. курсы генетики и механики развития), а понимание того факта, что процесс геномогенеза, как особая форма биологического развития (движения), является начальной фазой онтогенеза, которая переходит в другую его фазу и в другую форму развития — морфогенез. На протяжении преемственных рядов поколений (онтогенезов), под творческим влиянием отбора, происходит историческая трансформация органических форм — их филогенез. Таким образом, все три, переходящие друг в друга, формы движения составляют взаимно-связанные звенья одной общей цепи развития. Из сказанного выше видно, что процессы наследственной изменчивости имеют большое эволюционное значение, почему мы и должны обратиться к рассмотрению этого вопроса.
Общие данные о значении наследственности и изменчивости в эволюционном процессе
а. Общее значение наследственности в эволюционном процессе. Наследственность имеет большое значение, как фактор, обеспечивающий преемственность последовательных поколений. Наследственные изменения закрепляются и комбинируются в потомстве. Без этой функции наследственности накопляющее действие отбора, а следовательно, и возникновение материала для развития исторически целостных структур, было бы невозможно.
Однако, эволюционное значение наследственности не должно быть сложно понято и преувеличено. Выше было указано, что нужно различать генотипическую и фенотипическую преемственность. Тем более нужно различать филогенетическую преемственность. В последнее время необходимость различать различные формы преемственности удачно была подчеркнута Кремянским (1941). Генотипическая преемственность, или наследственность в тесном смысле слова, обеспечивает, конечно, ход развития каждой отдельной особи; она обеспечивает индивидуализированное (вспомним, что наследственность и изменчивость суть равно необходимые стороны процесса наследования) воспроизведение генетических качеств в конкретных последовательных поколениях, а также — их комбинирование, т. е. возникновение комбинативных форм — результатов геномогенеза. Однако, преемственность фенотипов на протяжении филогенетического существования вида и его истории покоится не на неизменной наследственной основе, а на действии естественного отбора, в процессе которого выживают носители всех тех геномов, которые осуществляются в конкретных, так сказать, «оправдавших» себя адаптивных морфо-физиологических признаках. Таким образом, одни и те же признаки, и — подчеркнем здесь — одни и те же корреляции, ведущие к этим признакам, могут стойко сохраняться на измененной наследственной основе на протяжении очень больших промежутков времени. Следовательно, генотипическая преемственность имеет значение в известной константности свойств ближайших поколений, но вовсе не составляет причины их преемственности на протяжении длительной истории вида. Мы, например, не имеем никакого права утверждать, что генотип лингулы (Lingula), известной с силура и существующей поныне, идентичен ее силурийским генотипам, хотя морфологически современные формы сохранили силурийские черты организации. Последние обеспечены филогенетической преемственностью, а не генетической. Таким образом, вне теории отбора совершенно невозможно объяснить эволюционный процесс и широко распространенный факт константности определенных признаков.
В заключение отметим, что наследственность, как известно, закрепляет достигнутое многообразие потомства. Она обеспечивает, в частности, явление расщепления в потомстве. Если один из родителей был мутантом, обладающим новыми полезными признаками, то эти последние, как правило, не нивеллируются при скрещиваниях. Правда, рецессивные признаки не развиваются. Однако, они проявляются в F2 при расщеплении потомства.
Этот факт имеет огромное значение, впервые подчеркнутое К. А. Тимирязевым (1888). Он указал, что закон расщепления «имеет, очевидно, громадное значение для эволюции организмов, так как показывает, что скрещивание вновь появившихся форм не грозит им уничтожением, а представляет для естественного отбора широкий выбор между чистыми и смешанными формами». Это значение расщепления, как мы видели, предвидел и подчеркивал Дарвин, а Мендель подтвердил его своими точными опытами, в чем и состоит его основная заслуга.
Стало быть, мутант, обладающий полезными признаками и влившийся в популяцию через скрещивание с другими формами данного вида, не может быть поглощён самим процессом скрещивания. Его рецессивность (как правило, новые мутации рецессивны) не является угрозой его будущему. Законы наследственности, сами по себе, не препятствуют, а напротив, способствуют его проявлению в последующих поколениях.
б. Эволюционное значение различных форм изменчивости. Вообще говоря, изменчивость приводит к возникновению новых особенностей, к увеличению разнородности популяции вида. Однако, конечные результаты изменчивости определяются характером ее.
Результаты мутаций и геномогенеза наследственно закрепляются, т. е. воспроизводятся в последующих поколениях. Морфологическое и физиологическое выражение наследственных изменений далеко неодинаково. Интенсивный геномогенез и крупные мутации, как правило, вредны. Они нарушают установившиеся корреляции, всю систему организма и в конкретных условиях среды могут обусловить летальный исход. Ряд летальных мутаций был описан для дрозофилы. Ввиду этого сложился взгляд, что крупные наследственные изменения, в частности крупные мутации, не имеют большого эволюционного значения.
Основное значение в эволюции, как это и отмечал Дарвин, имеют мелкие индивидуальные изменения, в том числе и ускользающие от нашего наблюдения. Эти слабо выраженные результаты идущих из поколения в поколения мутаций и геномогенеза не вызывают коренных изменений в геноме и не имеют выраженного вредного значения. Именно этот факт и играет большую роль в эволюции. В свое время Тимирязевым, а затем Шмальгаузеном (1940) было подчеркнуто, что естественный отбор происходит не через выбор особей (как при искусственном отборе), а через истребление менее соответствующих условиям среды. Носители крупных мутаций, по указанным выше причинам, как правило, уничтожаются. Напротив, носители мелких наследственных изменений, не очень вредных (полезных, индиферентных), механически сохраняются. Следовательно, мелкие мутации могут свободно накапливаться в популяции вида. Однако, и мелкие наследственные изменения не означают эволюции. Они только повышают разнородность населения вида и не имеют, сами по себе, эволюционного значения. Напротив, было указано, что всякое бесконтрольное суммирование мутаций приводит, в конечном счете, к распаду механизма индивидуального развития и дезорганизации (Шмальгаузен, 1940). Подобные явления мы наблюдаем, однако, преимущественно у культурных форм, т. е. у форм, которые, в той или иной степени, свободны от действия естественного отбора. Напротив, в естественных условиях такая дезорганизация, как правило, не наблюдается. Эти факты показывают, что и накопление мелких наследственных изменений, само по себе, не означает эволюции. Они с очевидностью приобретают эволюционное значение только под контролем отбора, и сами по себе должны рассматриваться лишь как основной материал для него.
Вопрос осложняется, далее, тем, что мутация только тогда может фактически влиться в популяцию вида, если ее носитель (мутант) скрестится с другими особями, или если мутанты размножаются. В результате скрещивания открывается путь к возникновению комбинативной изменчивости.
Эволюционное значение комбинативной изменчивости (геномогенеза) очень велико и, в известном смысле, последняя играет более значительную роль, чем мутации. Последние являются первичными наследственными изменениями. Так как мутанты (по крайней мере у форм, размножающихся путем копуляции разноименных гамет) вливаются в популяцию через скрещивание, то в результате последнего возникают комбинативные формы — продукт геномогенеза. Естественный отбор и имеет дело фактически не с мутантами, а с комбинативными формами, обладающими комбинированными признаками. Легко понять, что комбинации оказываются в различной степени удачными, жизнеспособными. Именно в их среде и происходит естественный отбор, который оказывается, таким образом, фактором, уничтожающим менее приспособленные комбинативные формы и в конечном счете строющим и создающим наиболее приспособленные комбинативные формы. Ни мутанты, ни комбинативные формы еще не означают эволюции, оставаясь лишь материалом для нее.
Обратимся теперь к эволюционному значению модификаций. Модификации являются формой проявления конкретных ненаследственных изменений в различных условиях среды.
Естественно ожидать, что если, например, новая мутация (в тех же условиях среды) выразится в новой модификации, то последняя не будет обязательно полезной. Она может быть и вредной. В процессе отбора сохраняются, однако, формы, модификационные изменения которых либо индиференты, либо полезны. При этом модификации выражаются в развитии определенных признаков, или морфозов. Полезные модификации (и морфозы) должны быть названы адаптивными, так как через конкретные адаптивные модификации должен происходить отбор конкретных геномов. Отбор, прежде всего, имеет дело с конкретными признаками. Отсюда ясно, что адаптивные модификации, выраженные в адаптивных морфозах, должны иметь очень большое эволюционное значение. В самом деле, всякая модификация есть реакция конкретного генома на определенные изменения условий среды. Следовательно, если данная модификация полезна, то через выживание ее носителей должно быть достигнуто выживание носителей соответственных геномов.
Таково общее эволюционное значение модификаций. Однако, в последнее время были выяснены факты, которые значительно углубили проблему эволюционного значения модификаций.
Было выяснено, прежде всего, что те модификации, которые мы обычно наблюдаем у организмов, не исчерпывают их модификационных возможностей. При воздействии на организм какими-либо новыми факторами, у организмов проявляются новые, ранее не наблюдавшиеся модификационные изменения. Следовательно, модификационная изменчивость организмов, по своим возможностям, оказывается шире, чем это требуется данными условиями среды. Мы, например, видели, что африканская антилопа-гну, перевезенная в суровые условия зимы северного полушария, развивает густой зимний волосяной покров, хотя в условиях ее родины предпосылок к этому нет. С другой стороны, оленебык не обладает способностью к такой модификации. Легка понять, что в суровых условиях зимы данная модификация адаптивна. Наличие таких модификаций полезного значения составляет как бы «адаптивный фонд» (Поляков, 1936) организмов.
Понятно, что наличие такого «адаптивного фонда» имеет огромное эволюционное значение, так как создает большое количество шансов на выживание при изменениях среды и, следовательно, может удлинить геологическое существование вида, или будет способствовать расширению его ареала как в горизонтальном, так и вертикальном направлениях.
Например, тигр (Tigris tigris), обладая модификационной способностью приобретать на холоду зимний волосяной покров, занял широкий ареал от Суматры, Явы и Индии до сурового Амурского края. Последствием такого расширения ареала была энергичная внутривидовая диференцировка на местные географические расы, а мы уже знаем, что этот процесс ведет к обособлению новых видов и, следовательно, к усилению темпов эволюции. Например, в Амурском крае образовался особый подвид тигра Tigris tigris longipilis — длинноволосый амурский тигр.
Такова одна сторона эволюционного значения модификаций. Другая сторона вопроса связана с тем, что под покровом адаптивных модификаций вырабатываются однозначные с ними, т. е. фенотипически сходно проявляющиеся генотипические изменения. Нужно, прежде всего, уточнить различия между теми и другими. Например, зимний волосяной покров в одних случаях проявляется только тогда, когда животное подвергается воздействию холода. Если его нет, то зимний волосяной покров не развивается. В этих случаях развитие зимнего волосяного покрова будет следствием и выражением модификации, так как он оказался прямым результатом, или прямой реакцией на холод (пример — антилопа-гну). Однако, известно много случаев, когда зимний волосяной покров развивается и без наступления холода, но примерно к тому времени, когда в данной области, как правило, наступают холода. Если, например, содержать обыкновенную европейскую белку в условиях постоянной комнатной температуры, то с наступлением зимы у нее и в комнатных условиях развивается светлый и густой зимний мех. Это наблюдается вообще у всех пушных зверей северного полушария. Совершенно ясно, что такое утепление меха непосредственно не связано с воздействием холода и не является модификацией. Мы имеем здесь дело с наследственным ритмом сезонной изменчивости (Ильин, 1926). Появление зимнего волосяного покрова и способность к сезонному ритму имеет здесь мутационное происхождение.
Таким образом, зимний волосяной покров может быть результатом модификаций и результатом мутаций. В этом смысле выше и говорилось об однозначных, параллельных модификациях и мутациях. Мы знаем в настоящее время много примеров такой однозначности или параллелизма тех и других. Таковы модификационный и генотипический меланизм у бабочек; модификационное укорочение ушей и хвоста у мышей и крыс под влиянием пониженной температуры и наследственно обусловленная укороченность тех же придатков у северных форм (песец, тобольская лисица); развитие мозолей под влиянием трения (модификация) и наследственно обусловленное появление их уже у зародышей, на тех местах, где трение еще только будет иметь место по достижении известного возраста (например, на коленных суставах бородавочника); сильная мускулатура, явившаяся результатом упражнения (модификационные изменения) и прирожденная; удлинение задних конечностей у животных с ампутированными передними ногами (модификационное изменение) и прирожденные длинные задние ноги при коротких передних (кенгуру, тушканчик) и т. д.
Чем объясняется этот параллелизм модификаций и мутаций? Можно было бы думать, что изложенные факты объясняются следующим образом. Модификационные признаки закрепляются во времени и становятся наследственными. Например, мозоли возникли, как реакция на рабочее трение, а затем стали наследственными.
Напомним, однако, что наследственны не признаки сами по себе. Поэтому самая постановка проблемы наследственного закрепления признаков неверна. Такой проблемы, строго говоря, не существует. Мало того, принимая идею наследственного закрепления признаков, мы становимся на почву метафизики. Например, хорда, если ее рассматривать, как наследственный признак, существует, как таковая, с кембрия, и следовательно, надо допустить, что с кембрия существует неизмененное наследственное основание. В действительности, как это дальше разъясняется, хорда существует с кембрия по другим причинам, так как от кембрийских геномов не осталось, конечно, и следа.
Было предложено другое объяснение параллелизма мутаций и модификаций, заключающееся в том, что определенные адаптивные модификации исторически заменяются им однозначными мутациями.
Рассмотрим соответствующий пример.
Гаузе (1940) было показано, что если содержать отдельную линию инфузории Euplotes в среде с повышенной соленостью (от 2,5 до 5%), то размеры инфузорий уменьшаются. Это уменьшение размеров является следствием модификации, имеющей физиологическое приспособительное значение. Далее, Гаузе показал, что в смешанной популяции тон же инфузории наблюдаются формы, наследственно маленькие. Если такую смешанную популяцию, содержащую «модификационно маленьких» и «наследственно маленьких» инфузорий воспитывать в водной среде с повышенной соленостью, то выживают «наследственно маленькие» формы, а не «модификационно маленькие». Таким образом, мутант оказался жизнеспособнее, чем параллельный модификант, следовательно, произошла замена модификации мутацией.
Возникает вопрос, почему именно мутационное возникновение определенных полезных признаков оказывается выгоднее, чем модификационное?
Ответ на этот вопрос может быть различным. Например, Лукин (1942) полагает, что причина сказанного заключается в относительной независимости мутационно возникшего признака от колебаний факторов внешней среды. Далее было предположено, что при относительна устойчивых условиях среды мутационно возникшие изменения отличаются большей специализацией по сравнению с ненаследственной изменчивостью, т. е. ближе соответствуют условиям среды.
Нужно думать, что рассматриваемая проблема может быть поставлена следующим образом. Строго говоря, если возникла мутация, параллельная адаптивной модификации, то это еще не означает преимущества мутанта перед модификантом. Мы видели, что мутации, сами по себе, едва ли полезны, или даже вредны. Следовательно, преимущества мутанта перед ему параллельным модификантом теоретически более чем сомнительно. Простое появление параллельной мутации вовсе не предрешает процесса, и, следовательно, нельзя говорить о простой «замене» модификации параллельной мутацией. Но дело в том, что если появилась мутация, параллельная адаптивной модификации, то соответственный мутант, через скрещивание, включается в популяцию вида. В конечном счете образуется новая — назовем ее пока так — комбинативная разновидность, в наследственное основание которой включена новая мутация. Если эта разновидность выжила, т. е. прошла контроль отбора, то это случилось не только потому, что она обладает новой мутацией, но и потому, что она обладает и другими преимуществами, в комбинации с которым новая мутация и приобрела адаптивное значение. Таким образом происходит не простая «замена» модификации ей параллельной мутацией, а замена одной разновидности другой, носителем новой мутации. Новая разновидность вытесняет прежние формы (у которых то же свойство было только модификационным) не потому, что мутация «лучше» параллельной ей модификации, а потому, что новая разновидность всестороннее приспособлена к условиям среды, чем прежняя разновидность, у которой данное свойство было только модификационным.
Для пояснения рассмотрим следующий пример. Положим, что в условиях леса у данного вида имелась модификационная способность приобретать при высокой влажности темную окраску, соответствующую окраске лесной подстилки. Данная модификация полезна, так как она имеет значение покровительственной окраски. Положим далее, что появился мутант, у которого темная окраска наследственно обусловлена. Такой мутант не будет зависеть в развитии своей темной окраски от колебаний влажности, и следовательно при любом значении ее будет иметь покровительственную окраску. Однако, в других отношениях, как это и наблюдается, данная мутация может быть вредной. Например, она может быть связана с пониженной жизнеспособностью. Таким образом, сама мутация еще не дает преимущественности. Однако мутант включается через скрещивание в популяцию. Положим, далее, что появились новые мутанты, которые характеризуются, например, новым поведением: при опасности они вжимаются в грунт и принимают неподвижную «позу покоя», вследствие чего становятся менее заметными. Такие мутанты также влились в популяцию. В конечном счете возникают новые формы в различных комбинациях. Преимущество получит та из них, которая обладает: 1) высокой жизнеспособностью, 2) наследственно обусловленной покровительственной окраской и 3) наследственно обусловленным поведением, выраженным в позе покоя во время опасности.
Сформировалась, следовательно, разновидность, явно преимущественная по сравнению с той, у которой способность темнеть была только модификацией. Произошла «замена» модификации ей параллельной мутацией, а по существу — замена одной (старой) разновидности новой, у которой наследственно темный цвет скомбинирован с рядом других полезных Свойств. Именно эта разносторонняя приспособленность новой разновидности, как целостного организма, и является причиной исторической «замены» модификации мутацией.
Таково объяснение явлений параллелизма модификаций и мутаций. Оно удовлетворительно освещает это явление и показывает большое эволюционное значение модификаций, состоящее в том, что под «покровом» адаптивных модификаций вырабатываются параллельные им приспособительные мутации, связанные, через скрещивание, с более всесторонней приспособленностью к среде и повышенной выживаемостью. Итак, процесс замены модификации ей параллельной мутацией протекает, как процесс смены одних разновидностей другими, где преимущество новых разновидностей выражается, как правило, в часто неуловимых, тонких, генетико-физиологических изменениях.
Из изложенного материала видно, что эволюционное значение различных форм изменчивости и процесса наследования выясняется только в связях этих процессов с естественным отбором. Процесс наследственных изменений и процесс наследования в целом протекают относительно быстро, и во всяком случае быстрее эволюции самого вида. Таким образом, при сведении эволюции к изменчивости, а константности видовых признаков — к наследственности, нельзя объяснить ни эволюции вида, ни относительной константности видовой формы. Наследственность, как генетическая преемственность, не объясняет известного постоянства видовых признаков. Геномы изменяются в рядах последовательных поколений, а видовые признаки сохраняются на протяжении геологического времени. Совершенно очевидно, что и изменения видовой формы и явления ее константности могут быть объяснены, если мы учтем еще один фактор, имеющий решающее значение. Этим фактором и является естественный отбор.